
焼き肉屋さんにいくとホルモンの一種としてウシの胃(前胃)であるミノ・ハチノス・センマイ・ギアラという4つの部位を見かけることがあります。
癖があることに加え内蔵の一種なので、好き嫌いが別れますが、コリコリとした食感でお酒にも合うので好んで食べる人もいるでしょう。
食べるだけだとあまりその正体や役割については考えが及ばないかもしれませんが、本来はウシを含む反芻動物にとって重要な役割を持っています。
畜産に関わる人にとっては必須の知識と言ますし、一般の人も常識として知っておくのも悪くないでしょう。
ただ、反芻動物の胃は呼称がやたらめったら多く、かつ機能と形の対応関係が分かりにくいと感じることが多いです。
しかし、名称の多さはともかく、形態と機能と順番に関してはよく考察してみると必然が高いことが分かります。
そこでここでは反芻動物の胃の名称と形態の簡単な覚え方をまとめておきます。
簡潔にまとめるとミノ(発酵)→ハチノス(分別)→センマイ(吸収)→ギアラ(消化)となっています。
反芻とは?反芻を行ったり前胃をもつ理由は?
反芻とは一度飲み込んだ草を再び口に戻して咀嚼する行為です。
ウシを観察していると長期に渡ってずっと口をもぐもぐしている様子が観察できますが、これは一度飲み込んだものを胃が吐き戻してもう一度噛んでいるのです。
人間にとっては少々気持ち悪い行為ですが、これは草食性の反芻動物を草地において圧倒的に有利にする適応なのです。
吐き戻した草はそのまま口に戻っているわけではありません。反芻動物の胃は4つに分かれており、その一番前の1つ目の胃(第1胃)で微生物を飼っており、一度、餌の草を発酵させています。なお、吐き戻された草に胃酸はかかっていません。
草の中にあるセルロースやヘミセルロースなどの多糖類を含む植物細胞壁は非常に丈夫で、本来は通常の動物の消化能力ではわずかしか消化できないものですが、反芻動物は発酵によって揮発性脂肪酸(VFA、酢酸・プロピオン酸・酪酸など)を作りこれが栄養源となります(Mackie, 2002; Bao et al., 2019)。
ただし大きな草のままでは微生物も流石に発酵がうまくできません。そこで反芻動物は繰り返し口に戻すことによって歯で物理的に破壊し、微生物の発酵を手助けしているのです(Matsuda et al., 2011)。
これによって他の哺乳類を凌駕して少ない草からでも効率的に栄養を補給することができます(Hofmann, 1989)。
しかもそれだけではなく微生物自体も微生物自身のためにタンパク質を合成しますが、反芻動物は餌中の尿素と肝臓で合成された尿素を第1胃に送り、微生物にタンパク質を合成させ、反芻動物はこれも飲み込んでしまうことで自分のタンパク質にしてしまいます(Hailemariam et al., 2021)。
更に胃の微生物は一部の植物二次代謝物(フェノール類・アルカロイド等)を分解でき、これが雑食的・採食幅を広げる(耐毒性を高める)助けになることも指摘されています(Loh et al., 2020)。
このように非常に効率的な機能に思えますが、これは草食に完全に特化した機能といえ、果実食・肉食・雑食の動物にとっては柔軟性がなくこの豪華な胃を維持する採算がとれません(トレードオフ)。そのためヒトやブタがこのような胃を持つことはないのです。
反芻動物の種類は?
学術的には4つの胃(四室胃)からなる「真の反芻動物(Ruminantia)」は鯨偶蹄目の反芻亜目に含まれる動物のことを指し、反芻亜目はマメジカ下目と真反芻下目の2つの下位群と6科に分かれ、現生種はおよそ200種程度とされます(Bao et al., 2019)。主要な科と代表例は次の通りです。
- ウシ科 Bovidae:ウシ属・ヒツジ属・ヤギ属・カモシカ属・アンテロープ類など。
- シカ科 Cervidae:シカ属(ニホンジカ)・ヘラジカ属・トナカイ属など。
- キリン科 Giraffidae:キリン・オカピ。
- プロングホーン科 Antilocapridae:プロングホーン。
- ジャコウジカ科 Moschidae:ジャコウジカ属など。
- マメジカ科 Tragulidae:マメジカ属など。
ただし、四室胃ではないものの、「反芻(chewing the cud)」という行動や「前胃発酵(foregut fermentation)」を行う動物は他にも居ます。これらは別の系統から収斂進化したものなので通常「反芻動物」とはされません。反芻動物に比べると単純な胃なので草からの栄養吸収効率も低くなっています。
ラクダ科(Camelidae:ラクダ、ラマ、アルパカ等)は鯨偶蹄目ではありますが、核脚亜目(ラクダ亜目)に属し、広く「反芻に似る」消化をしますが、胃は3室からなり、反芻動物における第3胃(葉胃)の吸収にあたる機能がありません(Fowler, 2008)。反芻亜目とは別に進化しました。
カバ科も鯨偶蹄目ではありますが、鯨河馬形類というクジラ類に近い仲間で、反芻は行わないものの、前胃(forestomach)で微生物発酵を行う前胃発酵を行います(Clauss at al., 2004)。
多くの大形有袋類(例:アカカンガルー、オーストラリアの大型マクロポッド)は前胃で前胃発酵を行いますが、通常の意味での「反芻(規則的な反復的再咀嚼)」は行いません(Vendl et al., 2017)。しかし、吐き出しと再咀嚼を示す個体記録があり、これを偽反芻(pseudo-rumination)またはメリシズム(merycism)と呼んでいます。メリシズムは反芻よりもはるかに短く、一貫性もないので、反芻とは区別することが多いです。
テングザル Nasalis larvatus も葉や果実を摂取する仲間で、メリシズムと前胃発酵を行います(Matsuda et al., 2011)。
ツメバケイ Opisthocomus hoazin は鳥ですが拡大した前胃(crop)で強い微生物発酵を行う非常に特殊な例で、化学的な発酵産物(揮発性脂肪酸)や微生物群集が哺乳類の前胃発酵者に類似していますが反芻はしません(Grajal, 1995)。餌をある程度分解・発酵させた物質を子へ与えることは報告されています。
反芻動物の胃(前胃)の命名と役割の一覧
以下に反芻動物の胃の順番からの命名、見た目からの命名、焼き肉での命名、役割について表にしてまとめておきます。
第1胃から第4胃にかけて発酵→分別→吸収→消化を行っていることが分かります。
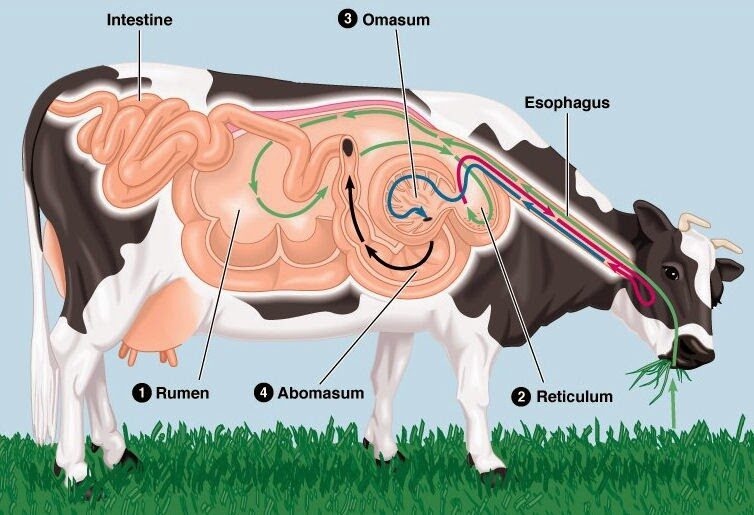
| 順番からの命名 | 見た目からの命名 | 焼き肉での命名 | 英名 | 役割 |
| 第1胃 | 瘤胃 | ミノ(蓑) | ルーメン(rumen) | 他の胃に比べてもコブ状に顕著に肥大し、乳頭に生息した微生物がセルロースや澱粉を発酵し、揮発性脂肪酸を産生。 |
| 第2胃 | 網胃・蜂巣胃 | ハチノス(蜂の巣) | レティキュラム(reticulum) | 網状の梁で小粒子・液相を選別して次の通路へ送り、逆に大きな粒子を送り返して反芻(再咀嚼)を促進。 |
| 第3胃 | 葉胃・重弁胃 | センマイ(千枚) | オマソン(omasum) | 多数の葉から水・電解質・一部の揮発性脂肪酸を吸収し、消化物の固形分比を高めて第4胃へ輸送。 |
| 第4胃 | 皺胃・真正胃 | ギアラ(偽腹)・ギャラ・赤センマイ・アボミ | アボマスム(abomasum) | 腺性粘膜を持ち消化を担う「真の胃」で、塩酸やペプシンを分泌し、微生物タンパクや食物タンパクを酸加水分解。 |
第1胃(瘤胃・ミノ・ルーメン)の名前の覚え方と役割は?

第1胃は見た目の命名としては瘤胃、焼き肉ではミノ、英名ではルーメンと呼ばれています。
第1胃の最大の特徴は他の胃に比べてもコブ状に顕著に肥大していることで、このことから瘤胃と呼ばれています。
ミノという焼き肉での名前は開いた様子が「蓑」に似ているためです。
この形態は第1胃の役割と深く関係しています。
第1胃は微生物(細菌・原虫・古細菌・菌類)がセルロースや澱粉を発酵し、揮発性脂肪酸(VFA;酢酸・プロピオン酸・酪酸)を産生します。VFAが主要なエネルギー源になり、微生物はタンパク質やビタミンも供給します(Perez et al., 2024)。運動(攪拌)とゲップ(eructation)でルーメンガスを排出します。
つまり第1胃は発酵を促す微生物の生息場所を広げるために「コブ」や「蓑」のように特別に肥大化させているのだと考えられるでしょう。
更に第1胃の内面には多くの乳頭(papillae)が発達して表面積を増やしVFA吸収を助けています(Pokhrel & Jiang, 2024)。
英語のルーメン rumen という呼称は、ラテン語の rūmen から借用されたものでこの語源は諸説ありますが、祖イタリック語 *roug(s)mən ~ *rug(s)mən < インド・ヨーロッパ祖語 *h₁rewg-, *h₁rewǵ- 「げっぷをする」に由来すると考えられています。
この名称もよく第一胃の役割を表していて、この第一胃では主にメタン生成古細菌(methanogenic archaea) による水素(H₂)やメチル化化合物の還元でメタンが生成され、二酸化炭素(CO₂)45~75% と メタン(CH₄)20~35% を主成分とするルーメンガスが発生し、反芻動物はゲップでガスを排出します(Zhang et al., 2020)。
温室効果ガスとしてウシのメタンを含むゲップが問題視されることがありますが、この原因は第1胃によるものと言えます。
第1胃はコブ状に多くなった蓑で微生物を飼い、不要なゲップを作る場所だと考えれば覚えやすいです。
なお上述のように微生物によってタンパク質の合成や植物二次代謝物(フェノール類・アルカロイド等)の解毒も行われています。
第2胃(網胃・ハチノス・レティキュラム)の名前の覚え方と役割は?

第2胃は見た目の命名としては網胃、焼き肉ではハチノス、英名ではレティキュラムと呼ばれています。
網胃やハチノスという呼称は胃の内壁の粘膜が蜂の巣のように六角形のひだ状になっていることに由来します。
英語のレティキュラム reticulum はラテン語 rēticulum 「網」から借用され、ラテン語 rēte 「ネット、スネア」+ -culum (指小辞) < イタリック祖語 *rēti- < インド・ヨーロッパ祖語 *h₁reh₁- 「別々の、ゆるい」に由来します。英語で銃のスコープや顕微鏡の照準線を意味するレティクル reticle と同源です。
つまりどれも多角形の網状の構造があることに由来しています。
この形態も第2胃の役割と深く関係しています。
第2胃は収縮して小粒子・液相を選別して次の通路へ送り、逆に大きな粒子を送り返して反芻(再咀嚼)させます(Song et al, 2024)。ルーメンと網胃は連結して機能し、総称して「網ルーメン(reticulorumen)」または「反芻胃(ruminoreticulum)」とも呼ばれます。
この選別のときに蜂の巣状または網状と呼ばれる梁(pillars)が作り出す多室的な空間配列が「浅い籠」のようになり、流速と選別効率を制御します。
つまり大型の粒子は籠の内部にとどまりもう一度第1胃に繰り返され、小粒子や液体は第3胃以降に流れていきます。逆に消化機能はわずかしかありません。
この特徴から金属などの異物が留まりやすくウシでは特に創傷性網胃腹膜炎(hardware disease)という病気になってしまうことがあります。
第2胃は蜂の巣のような網状構造で反芻する必要がある餌の選別を行う器官だと考えれば覚えやすいです。
第3胃(葉胃・センマイ・オマソン)の名前の覚え方と役割は?

第3胃は見た目の命名としては葉胃、焼き肉ではセンマイ、英名ではオマソンと呼ばれています。
葉胃という呼称は葉っぱや弁のような形をしたヒダ(薄い板状の組織)が何枚も幾重にも重なり合っている構造をしていることに由来します。重弁胃も同様です。
センマイという呼称は内部に無数のヒダが重なり合って「千枚」あるように見える外観に由来してるともされます。韓国語の「チョニョブ(千葉・千枚)」の言葉が由来であるという説もありますが、概ね意味は同じです。
つまりほとんど同じ特徴に由来しています。
この形態も第3胃の役割と深く関係しています。
第3胃は水・電解質・一部のVFAを吸収し、消化物の固形分比を高めて第4胃へ送ります。液体の除去とさらなる微小粒子の分離を行っていると言えます(Pérez et al., 2023)。
このとき多数の葉(laminae)によって大きな吸収表面を作ることで効率的な水分・VFA吸収を実現しています。
第3胃は千枚にも見える無数の葉状のヒダで腸に先んじて液状部分の吸収を行っていると考えれば覚えやすいです。
なお、オマソン omasum はラテン語の omāsum「雄牛の胃袋」からの借用語で、ラテン語はガリア語由来し、ガリア語はカルタゴ語から借用、カルタゴ語はセム祖語の*ḥamṯ-「腹部」に由来すると考えられています。
オマソンに関しては原義から大きく意味が変わってしまっています。
第4胃(皺胃・ギアラ・アボマスム)の名前の覚え方と役割は?
第4胃は見た目の命名としては皺胃、焼き肉ではギアラ、英名ではアボマスムと呼ばれています。
皺胃という呼称は粘膜面に細かいヒダがあることに由来します。
真正胃という呼称は反芻動物以外の通常の胃と同じ機能を持っていることに由来します。
第4胃は腺性粘膜を持つ「真の胃」で、塩酸やペプシンを分泌し、微生物タンパクや食物タンパクを酸加水分解して小腸での吸収に備えます(Ash, 1961)。この意味で「真正」ということです。
更に第4胃にシワがあることにも意味があります。
折りたたまれた皺は、空のときは折りたたまれていて、充満時には伸びて胃腔容積を増やせます(可変体積)。また皺があることで胃壁の運動(攪拌・混和)が効率的になり、酸と内容物の接触面が増えて消化が進みやすくなります。これらは特別なわけではなく、ヒトを含む一般的な「胃のルガ(rugae)」の機能と一致します。
更に、粘膜が折りたたまれていることで塩酸やペプシンなどの胃液を分泌する胃腺を収める面積が増え、酸・酵素の分泌や粘液分泌が効率よく行えます。
したがって、第4胃は他の哺乳類と同じタンパク質の化学消化機能を持つ「真の胃」で、シワ状のヒダに胃腺を持ち、食物の一時的に蓄える機能があると考えれば覚えやすいです。
アボマスム abomasum は新ラテン語 abomāsum < ab- 「〜から離れて」 + omāsum 「雄牛の胃袋」に由来します。つまり「オマソン(第3胃)から離れた胃」という意味です。
なお、ギアラという呼称ですが、由来は胃でありながらも腸のような働きをすることから、偽の腹という意味の「偽腹(ぎばら)」が訛なまったとする説がインターネットでは流布されています。
しかし、腸のような役割を持っているのは第3胃で、第4胃は上述のように胃そのものなので、この由来説はあまり納得できません。第3胃と混同したということならまだ分かりますが、その証拠はありません。
他には米軍基地で働いていた人が報酬(ギャランティー)としてもらっていたことから「ギャラ」→「ギアラ」になったという説もありますが(田辺,2016)、歴史学的な経緯は不明でこれも怪しいものがあります。
引用文献
Ash, R. W. 1961. Acid secretion by the abomasum and its relation to the flow of food material in the sheep. The Journal of Physiology 156(1): 93-111. https://doi.org/10.1113/jphysiol.1961.sp006660
Bao, W., Lei, C., & Wen, W. 2019. Genomic insights into ruminant evolution: from past to future prospects. Zoological Research 40(6): 476-487. https://doi.org/10.24272/j.issn.2095-8137.2019.061
Clauss, M., Schwarm, A., Ortmann, S., Alber, D., Flach, E. J., Kühne, R., … & Hofer, H. 2004. Intake, ingesta retention, particle size distribution and digestibility in the hippopotamidae. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 139(4): 449-459. https://doi.org/10.1016/j.cbpb.2004.10.002
Fowler, M. E. 2008. Camelids are not ruminants. Zoo and Wild Animal Medicine (Sixth Edition), pp.375-385. ISBN: 9781416057598, https://doi.org/10.1016/B978-141604047-7.50049-X
Grajal, A. 1995. Structure and function of the digestive tract of the hoatzin (Opisthocomus hoazin): a folivorous bird with foregut fermentation. The Auk 112(1): 20-28. https://doi.org/10.2307/4088763
Hailemariam, S., Zhao, S., He, Y., & Wang, J. 2021. Urea transport and hydrolysis in the rumen: a review. Animal Nutrition 7(4): 989-996. https://doi.org/10.1016/j.aninu.2021.07.002
Hofmann, R. R. 1989. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system. Oecologia 78(4): 443-457. https://doi.org/10.1007/BF00378733
Loh, Z. H., Ouwerkerk, D., Klieve, A. V., Hungerford, N. L., & Fletcher, M. T. 2020. Toxin degradation by rumen microorganisms: a review. Toxins 12(10): 664. https://doi.org/10.3390/toxins12100664
Mackie, R. I. 2002. Mutualistic fermentative digestion in the gastrointestinal tract: diversity and evolution. Integrative and Comparative Biology 42(2): 319-326. https://doi.org/10.1093/icb/42.2.319
Matsuda, I., Murai, T., Clauss, M., Yamada, T., Tuuga, A., Bernard, H., & Higashi, S. 2011. Regurgitation and remastication in the foregut-fermenting proboscis monkey (Nasalis larvatus). Biology Letters 7(5): 786-789. https://doi.org/10.1098/rsbl.2011.0197
Perez, H. G., Stevenson, C. K., Lourenco, J. M., & Callaway, T. R. 2024. Understanding rumen microbiology: an overview. Encyclopedia 4(1): 148-157. https://doi.org/10.3390/encyclopedia4010013
Pérez, W., Duro, S., & Gündemir, O. 2023. Anatomical Differences in the Omasum of Weaning Calves Fed with Different Diets. Anatomia 2(2): 176-188. https://doi.org/10.3390/anatomia2020016
Pokhrel, B., & Jiang, H. 2024. Postnatal growth and development of the rumen: integrating physiological and molecular insights. Biology 13(4): 269. https://doi.org/10.3390/biology13040269
Song, Y., Lan, X., Liu, L., Wan, F., Shen, W., & Wang, Z. 2024. Exploring the reticulo-ruminal motility pattern in goats through medical barium meal imaging technology. Frontiers in Veterinary Science 11: 1371939. https://doi.org/10.3389/fvets.2024.1371939
田辺晋太郎. 2016. 牛肉論. ポプラ社, 東京. 169pp. ISBN: 9784591152461
Vendl, C., Munn, A., Leggett, K., & Clauss, M. 2017. Merycism in western grey (Macropus fuliginosus) and red kangaroos (Macropus rufus). Mammalian Biology 86: 21-26. https://doi.org/10.2307/4088763
Zhang, Z., Wang, Y., Si, X., Cao, Z., Li, S., & Yang, H. 2020. Rumen methanogenesis, rumen fermentation, and microbial community response to nitroethane, 2-nitroethanol, and 2-nitro-1-propanol: an in vitro study. Animals 10(3): 479. https://doi.org/10.3390/ani10030479

