
ミミガタテンナンショウ・ヒガンマムシグサ・マムシグサはテンナンショウ類の中でも代表的な種類です。テンナンショウ属は極めて形が類似しており、区別が難しいグループです。はっきりいってテンナンショウ属の区別は極めて困難です。分類に関しても地域個体群間で変異が大きく、どこまでを種とするかで頻繁に分類が変わっており、素人が判断するのは難しいでしょう。しかし、ミミガタテンナンショウ・ヒガンマムシグサ・マムシグサに限れば仏炎苞を観察することで区別が可能です。仏炎苞は非常に特殊化したサトイモ科固有の形態ですが、最近の研究では驚くべきことにこの内部にキノコバエ類を死ぬまで閉じ込めることで受粉を行っている事がわかっています。更に盛んに日本で行われている研究ではこのキノコバエ類の種類はテンナンショウの種類ごとに異なっており、競合や交雑を防いでいることも分かってきました。真っ赤に熟す果実についても一見テンナンショウ属で共通でよく似ており、鳥によって種子散布するという点は変わりませんが、一部の種類では果期が夏になっており、ヒヨドリに特化した種子散布を行っている可能性が示されています。本記事ではテンナンショウ属の分類・送粉生態・種子散布について解説していきます。
日本に局所的に分布する極めて類似したテンナンショウ類
ミミガタテンナンショウ(耳形天南星) Arisaema limbatum は日本の本州(東北地方~中部地方東部の太平洋側・兵庫県)・四国(高知県沖の島)・九州(大分県)のみに局所的分布し(邑田ら,2018)、落葉樹林や林縁に生える多年草です(邑田,2015)。
ヒガンマムシグサ(彼岸蝮草) Arisaema aequinoctiale は日本の本州(関東・中部地方・広島県・山口県)・四国のみに分布し、海岸近くの照葉樹林下などに生える多年草です。
マムシグサ(蝮草) Arisaema japonicum は日本の四国・九州(吐噶喇列島まで);韓国(鬱陵島)に分布し、低地から山地までの林下・林縁に生える多年草です。
いずれもサトイモ科テンナンショウ属で、極めて形が類似しています。この属の他の植物と異なる特徴的な共通構造としては、茎が地下茎となっており、扁球状の球茎があるということが挙げられます。この上に「偽茎」が生え、偽茎を巻くように「鞘状葉」が付いています。光合成を担うのは偽茎の先に生える「普通葉」で、掌状複葉、輪状複葉、稀に単葉になっています。複葉の場合、分かれた部分を「小葉」と呼びます。1個の葉に1個の腋芽をつけます。

テンナンショウ属は植物体にシュウ酸カルシウム結晶が含まれ哺乳類には毒性があることや、性転換することでも有名です(河野,1988)。
ミミガタテンナンショウ・ヒガンマムシグサ・マムシグサの違いは?
はっきりいってテンナンショウ属の区別は極めて困難です。分類に関しても地域個体群間で変異が大きく、どこまでを種とするかで頻繁に分類が変わっており、その度に書籍が出版されています(邑田, 2015;2018)。最新の文献では日本のテンナンショウ属は53種に分けられています(邑田,2018)。
したがって素人が簡単に区別するのは難しいでしょう。
ここでは、比較的著名なミミガタテンナンショウ・ヒガンマムシグサ・マムシグサに限って違いについて考えてみます。より詳しいことを知りたい人は是非書籍を読んでみてください。
これら3種はまず、ミミガタテンナンショウとヒガンマムシグサでは花序柄は(少なくとも雌では)偽茎より長く、小葉は5~7枚かそれ以上で、子房あたり平均胚珠数は10個以上であるのに対して、マムシグサでは花序柄は偽茎より短く、小葉は7枚以上で、子房あたり平均胚珠数は8個以下です。またマムシグサの仏炎苞の口部は耳状に広がりません。
ミミガタテンナンショウとヒガンマムシグサの違いとしては、ミミガタテンナンショウでは仏炎苞の口部が広く耳状に広がり、開出部の幅が8mm以上であるのに対して、ヒガンマムシグサでは仏炎苞の口部がやや耳状に広がり、開出部の幅が8mm未満であるという点が挙げられます。
また図鑑の検索表に書かれていませんが、ミミガタテンナンショウでは花序付属体が棒状からやや棍棒状で先端はふくれて円頭になり、仏炎苞口部から完全に外に出ますが、ヒガンマムシグサでは花序付属体が棒状で仏炎苞口部からやや出る程度という違いもあるでしょう。この点は比較的わかりやすいと考えています。
しかし、ミミガタテンナンショウとヒガンマムシグサはテンナンショウ属の中でも染色体数が2n=26であること、小葉が雌で9~17枚であることなど共通点が多く同定は難しい2種ではあります。
いずれにせよ、花が咲いていないと確実な同定は難しいです。

















テンナンショウ属の花の構造は?
テンナンショウ属は花の構造も他の植物とは大きく異なっており専門用語も多いです。
多くの種類では花は春から初夏に咲きます。
まずテンナンショウ属の花は「仏炎苞」と呼ばれる構造に包まれています。これはサトイモ科共通構造で、苞と呼ばれる葉が大型に発達したもので、内部に花序が見られます。
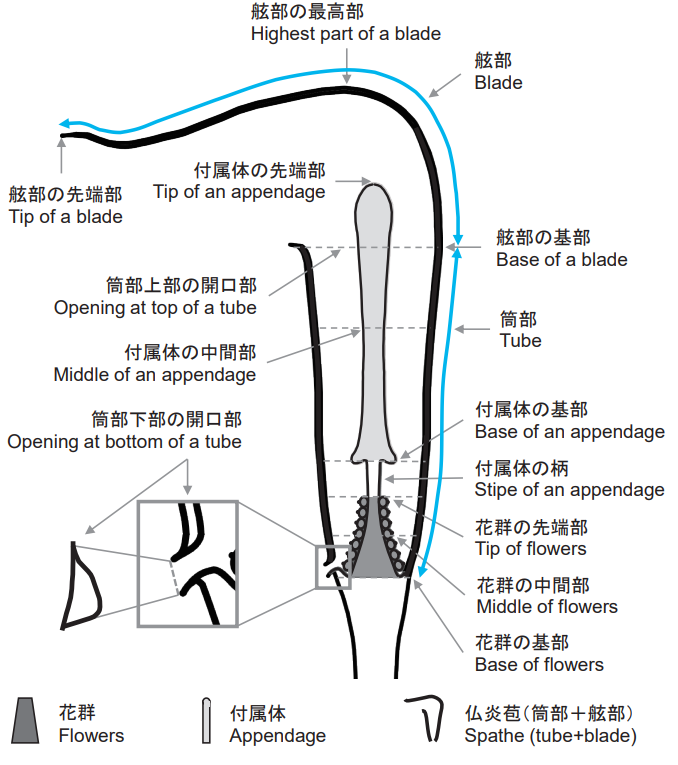
仏炎苞は「花序柄」で偽茎と繋がっており、構造としては花序を直接包んでいる「筒部」、花序への入り口に当たる「口辺部」、屋根状に口辺部を緩く塞いでいる「舷部」に分けられます。
サトイモ科の花序もまた特殊化しており、「肉穂花序」と呼ばれます。肉穂花序は花軸が肥厚して多肉質となり、その周囲に柄のない多数の花が密生しています。肉穂花序には「花序付属体」が付いており、上に伸びて、仏炎苞の筒部から確認できる場合があります。
テンナンショウ属は雌雄異株です。そのため、雄株と雌株があり、雄株には肉穂花序に雄花が、雌株には肉穂花序に雌花がつきます。
テンナンショウ属は興味深いことに栄養状態によって性別が決まります。小型株になった個体では、比較的資源が少なくても配偶子を作ることができる雄株に、栄養が足りて成長し大型株になった個体では資源を生かして雌花や両性の花を作る雌株に性転換します。このような特殊な雌雄別株は「雌雄偽異株」と呼ぶこともあります。
ミミガタテンナンショウは花期が暖地では3月~、東北地方では5月頃で、仏炎苞の口部が広く耳状に広がり、開出部の幅が8mm以上です。
ヒガンマムシグサは花期が3~4月で、仏炎苞の口部がやや耳状に広がり、開出部の幅が8mm未満です。
マムシグサは花期が3~4月で、仏炎苞の口部はやや開出する程度です。

テンナンショウ属の花にキノコバエは死ぬまで閉じ込められる!?
極めて特殊化しているテンナンショウ属の花ですが、どのような昆虫がやってきて受粉しているのでしょうか?非常に気になるところです。
このことは様々な研究者によって研究が行われてきました。その結果、なんとキノコバエ科 Mycetophilidae やクロバネキノコバエ科 Sciaridae といった非常に小型のハエの仲間がやってきていることがわかったのです(田中・平野,2000;Vogel & Martens, 2000;Suetsugu, 2022)。
これらのハエの仲間は幼虫の間にカビやキノコの菌糸などを食べるという生態を持ち、林内では沢山生息していることがよく知られています。テンナンショウ属が林内を生息地にすることを考えれば非常に良い送粉者であると言えるでしょう。
具体的にどのようにしてキノコバエは花にやってきてテンナンショウ属の花を受粉させるのでしょうか?
まずキノコバエの仲間は花序付属体を目印に仏炎苞に引き寄せられます。
残念ながらどのようにキノコバエの仲間が仏炎苞に引き寄せられるのかについてはまだよく分かっていません。ユキモチソウ Arisaema sikokianum ではキノコに擬態してキノコに似た匂いをだすため、この匂いに惹き寄せられてキノコバエの仲間は花にやってくると考えられていますが、他のテンナンショウ属ではこのような匂いとなる成分が現在のところ確認されていません。しかし、ヒトの鼻ではわからない、キノコバエの仲間の性フェロモンを出している可能性が指摘されています。
次に仏炎苞に引き寄せられたキノコバエの仲間は内部に侵入して、肉穂花序に到達します。
しかし、この仏炎苞の内側はつるつるしていて、更に鼠返しもついているので、匂いに引き寄せられたキノコバエは落ちて上から出られなくなってしまいます。そのためキノコバエの仲間は仏炎苞の内部で暴れまわることになります。
もし落ちたのが雄株の時は仏炎苞の下の方に出口があります。外に出る時には雄花の花粉を体に沢山つけさせられることになりますが、この時にはキノコバエの仲間が命に関わることはありません。
問題は雌株の時です。雌株の場合は雄株のように下に出口がありません!テンナンショウ属側は雄株の時と同様にキノコハエの仲間が仏炎苞の内部で暴れまわることによって受粉出来るのですが、キノコバエ側はそのまま死んでしまいます。
つまり、キノコバエの仲間にとっては2回も出られないように閉じ込められた上で、最後には殺されています。非常に残酷な受粉方法と言えるかもしれません。
なぜ殺してしまうのかについてはまだ十分に分かっているわけではありませんが、キノコバエの仲間が死ぬまで暴れ続けることが効率的な受粉に繋がっているという考えもあります。
テンナンショウ属の種類によって花に来るキノコバエが違っていた!?
確かにテンナンショウ属の受粉はユニークな方法ではありますが、不思議な点もあります。テンナンショウ属は多様化しており、同所的に別種が見られることもあります。このような場合、同じキノコバエの仲間を送粉者として利用していて、競合や交雑のような問題は発生しないのでしょうか?
この点に関連して日本で研究が行われています(Matsumoto et al., 2021)。この研究では岡山県大茅スキー場に分布するウメガシマテンナンショウ Arisaema maekawae・ホソバテンナンショウ Arisaema angustatum・コウライマムシグサ Arisaema peninsulae・ミヤママムシグサ Arisaema pseudoangustatum var. pseudoangustatum・ヒロハテンナンショウ Arisaema ovale の5種にやってくるキノコバエの具体的な種類が調べられました。
その結果、呼び寄せるキノコバエの種類はテンナンショウの種類ごとに異なっていることが分かったのです。
具体的にはウメガシマテンナンショウでは Megophthalmidia sp.、ホソバテンナンショウでは Cordyla sp.、コウライマムシグサではナミキノコバエ属の一種 Mycetophila sp.、ミヤママムシグサではTrichosia sp.、ヒロハテンナンショウでは Anatella sp. が最もやってくる割合が高くなっていました。
この事実はテンナンショウ類はキノコバエの中でも更に別の種類を花に惹き寄せることで、競合や交雑を防いでいると考えられます。
他にも香川県大滝大川県立自然公園でユキモチソウ Arisaema sikokianum とアオテンナンショウ Arisaema tosaense 間でもやはりやってくるキノコバエの種類が異なっていたことを示した研究や(Matsumoto et al., 2019)、奈良県天川村でホロテンナンショウ Arisaema cucullatum・キシダマムシグサ Arisaema kishidae・ムロウテンナンショウ Arisaema yamatense 間でもやってくるキノコバエの種類が異なっていたことを示した研究(Kakishima et al., 2020)、長野県北浅間山でヤマジノテンナンショウ Arisaema solenochlamis・コウライマムシグサ・ヤマトテンナンショウ Arisaema longilaminum・ユモトマムシグサ Arisaema nikoense subsp. nikoense var. nikoense 間でやってくるキノコバエの種類が異なっていたことを示した研究(前田・高橋,2021)、ナギヒロハテンナンショウ Arisaema nagiense とヒロハテンナンショウ Arisaema ovale 間でもやってくるキノコバエの種類が異なっていたことを示した研究もあります(Matsumoto et al., 2023)。
具体的にどのようなメカニズムによって異なったキノコバエの仲間がやってくるのかは分かっていません。しかし、上述のようにヒトの鼻ではわからない、キノコバエの仲間の性フェロモンを出しているのであれば、この成分の違いがこの違いを生み出しているのかもしれません。いずれにせよ、植物の種分化と昆虫の関係を示す興味深い結果であることは間違いないでしょう。
果実は液果で種子はおそらく鳥散布のみ?
果実はテンナンショウ属共通で液果で、卵球形です。果柄がないので集合果のように密集しています。熟すと緑色から朱赤色になり、少数の種子があります。種子はほぼ球形で乳白色、まれに斑点があります。
熟して赤くなることからも分かるように果実は明らかに動物に食べてもらえるようにアピールしており、種子散布は動物に頼っていることが窺えます。
しかしどのような動物でも食べるという訳ではなく、基本的には鳥によって果実は食べられ種子散布されます。日本の研究では哺乳類がこの果実を食べている様子は殆ど確認されておらず、唯一果実を食べるアカネズミは種子まで食害してしまうので種子散布には貢献しないと考えられます(大石ら,2020)。
哺乳類がテンナンショウ属の果実を食べない理由としては、果実にも哺乳類には有毒なシュウ酸カルシウム結晶が含まれているからだと考えられます。しかし、海外ではヒマラヤツキノワグマやアメリカグマが食べていた証拠があるので、ニホンツキノワグマが食べる可能性もあるという意見もあります(前田・高橋,2021)。
テンナンショウ属の果実は夏に生るグループと秋~冬に生るグループが居た!?
テンナンショウ属の果実も花序に生る数に違いはあるものの、基本的に種間でよく似ており、単体では区別出来ないほどです。しかし、テンナンショウ属の果期は興味深いことに2タイプあることが分かっています(小林,2017;小林ら,2017)。
テンナンショウ属の大部分である39グループについては果期が9月中旬~1月の秋~冬ですが、ヒガンマムシグサ群(ナガバマムシグサ Arisaema undulatifolium subsp. undulatifolium を除くタカハシテンナンショウ Arisaema nambae ・ハリママムシグサ Arisaema minus・ミミガタテンナンショウ・ヒガンマムシグサ・ウワジマテンナンショウ Arisaema undulatifolium ssp. uwajimense)の5グループとセッピコテンナンショウ Arisaema seppikoense の果期は6月下旬~8月の夏となっています。
このような生態の違いは、種子散布の担い手である鳥の種類にも違いをもたらしているのでしょうか?
このことを調べるために西日本(兵庫県・岡山県・広島県)で自動撮影カメラを用いた調査が行われています。
その結果、この調査で対象となった果期が秋~冬の7グループ(ホソバテンナンショウ・ムロウマムシグサ・アオオニテンナンショウ・ムロウテンナンショウ・ナンゴクウラシマソウ・ウラシマソウ・ムサシアブミ)ではシロハラ・ヤマドリ・ヒヨドリが果実を採食していました。また別の研究ですが南関東(神奈川県秦野)でカンヨウマムシグサを調べた結果はヒヨドリ・ルリビタキ・シロハラ・ヤマドリ・ソウシチョウが果実を採食することが確認されています。石川県の調査でもよく似た結果が示されています(大石ら,2020)。このうち、ヤマドリは体内で種子を破壊する種子食害者とされているので種子散布に貢献していない可能性が高いです。果実はゆっくり無くなっていきます。
一方、果期が6月下旬~8月の夏の4グループ(タカハシテンナンショウ・ハリママムシグサ・ミミガタテンナンショウ・セッピコテンナンショウ)ではヒヨドリとヤマドリが果実を採食するという異なる傾向が見られたのです。また別の研究の南関東でもヒヨドリのみがミミガタテンナンショウの果実を食べています。上述のようにヤマドリは種子散布に貢献していない可能性が高いので実質、ヒヨドリだけが種子散布を担っています。果実は急速に無くなっていきます。
つまり、果期が秋~冬のテンナンショウ類では多様な鳥が果実を食べるのに対して、果期が夏のテンナンショウ類ではヒヨドリだけと非常に限られた鳥のみが果実を食べていたということになります。

ではなぜこのような違いが見られたのでしょうか?
テンナンショウ属は果期が秋~冬が大多数であることから、元々、テンナンショウ属は果期が秋~冬だったのでしょう。
この時期は様々な渡り鳥が冬鳥として、餌となる果実を求めて日本列島にやってくる時期です。また漂鳥のように国内で南下する鳥も居ます。他の植物も同調するように果実をつけていきます。
そのため、テンナンショウ属は留鳥に加え、これらの冬鳥や漂鳥によって効率よく種子散布を行っていると考えられます。
しかし、一部のヒガンマムシグサ群やセッピコテンナンショウは夏に果実を作るように進化しました。その理由は完全にはまだ分かりませんが、ヒヨドリとの強い相互作用がその進化を促した可能性があります。
ヒヨドリは留鳥で、日本では他の鳥に比べても極めて沢山の個体数を持つ種類です。そのため、ヒヨドリが多い環境では、果実をつける植物種は少ない繁殖時期である夏(7~8月)に果実は不足気味になると考えられます。
そのような時期に、林床で遠目にもよく目立つ集合果をつけるテンナンショウ属の果実はヒヨドリの格好の餌になるのでしょう。
その結果、ヒヨドリに特化し夏に結実するグループが生まれ、ヒヨドリが殆どの果実を採食し、秋~冬に結実するグループよりも高い割合で果実が持ち去られることで、効率的に多様な環境に種子が散布されているのかもしれません。それが一部のヒガンマムシグサ群やセッピコテンナンショウだったのです。
また、これらの果期が夏のグループの雌株では花序柄が葉柄より長い傾向という傾向があります。この点も果実をヒヨドリに採食させるために花序柄を伸ばして、果序をアピールさせるために進化している可能性があります。
訪花昆虫のみならず、果期についても日本の動物と密接に関わりながら植物が変化しているという事実はこれらの生き物を切り離して考えるべきではないということを教えてくれるでしょう。
引用文献
Kakishima, S., Sueyoshi, M., & Okuyama, Y. 2020. Floral visitors of critically endangered Arisaema cucullatum (Araceae) endemic to Kinki region of Japan. Bulletin of the National Museum of Nature and Science. Series B, Botany 46(1): 47-53. https://www.kahaku.go.jp/albums/abm.php?d=4697&f=abm00001696.pdf&n=L_BNMNS_B46-1_47.pdf
小林禧樹. 2017. テンナンショウとともに30年―ヒガンマムシグサ群の調査・研究からみえてきたもの―. 分類 17(2): 113-127. https://doi.org/10.18942/bunrui.01702-02
小林禧樹・北村俊平・邑田仁. 2017. 日本産テンナンショウ属(サトイモ科)の果実熟期の分化と鳥類による種子散布. 植物研究雑誌 92(4): 199-213. https://doi.org/10.51033/jjapbot.92_4_10799
河野昭一. 1988. 植物の世界 ナチュラルヒストリーへの招待 第2号. 教育社, 東京. 143pp. ISBN: 9784315506631
前田夏樹・高橋一秋. 2021. テンナンショウ属の開花・結実・花粉媒介・種子散布―浅間山の事例. 長野大学紀要 42(3): 23-57. http://id.nii.ac.jp/1025/00001331/
Matsumoto, T. K., Miyazaki, Y., Sueyoshi, M., Senda, Y., Yamada, K., & Hirobe, M. 2019. Pre-pollination barriers between two sympatric Arisaema species in northern Shikoku Island, Japan. American Journal of Botany 106(12): 1612-1621. https://doi.org/10.1002/ajb2.1389
Matsumoto, T. K., Hirobe, M., Sueyoshi, M., & Miyazaki, Y. 2021. Selective pollination by fungus gnats potentially functions as an alternative reproductive isolation among five Arisaema species. Annals of Botany 127(5): 633-644. ISSN: 0305-7364, https://doi.org/10.1093/aob/mcaa204
Matsumoto, T. K., Sueyoshi, M., Sakata, S., Miyazaki, Y., & Hirobe, M. 2023. Two closely related species of the Arisaema ovale group (Araceae) selectively attract male fungus gnats of different Anatella species (Diptera: Mycetophilidae). Plant Systematics and Evolution 309(1): 4. https://doi.org/10.1007/s00606-022-01839-7
邑田仁. 2015. サトイモ科. In: 大橋広好・門田裕一・木原浩 (Eds.), 日本の野生植物 (改訂新版, Vol. 1, pp. 93-100). 平凡社. ISBN: 9784582535310
邑田仁・大野順一・小林禧樹・東馬哲雄. 2018. 日本産テンナンショウ属図鑑. 北隆館, 東京. 360pp. ISBN: 9784832610057
大石里歩子・前田大成・北村俊平. 2020. 日本の温帯林におけるサトイモ科カントウマムシグサの種子散布者としての鳥類の有効性: 果実の持ち去り量と発芽への影響. Bird Research 16: A1-A14. https://doi.org/10.11211/birdresearch.16.A1
Suetsugu, K. 2022. Arisaema: Pollination by lethal attraction. Plants, People, Planet 4(3): 196-200. https://doi.org/10.1002/ppp3.10261
田中肇・平野隆久. 2000. 花の顔―実を結ぶための知恵. 山と渓谷社, 東京. 191pp. ISBN: 9784635063043
Vogel, S., & Martens, J. 2000. A survey of the function of the lethal kettle traps of Arisaema (Araceae), with records of pollinating fungus gnats from Nepal. Botanical Journal of the Linnean Society 133(1): 61-100. ISSN: 0024-4074, https://doi.org/10.1006/bojl.1999.0317
出典元
本記事は以下書籍に収録されていたものを大幅に加筆したものです。

