ナスとワルナスビはどちらも「ナス」の名がつく名前と花の形が類似した2種ですので、見たことがない人は2種を混同するかもしれません。しかし、ナスとワルナスビは全く異なる種類です。ナスは栽培植物で自生することはなく刺がないですが、ワルナスビは野草として自生し刺があります。しかし、今でこそナスは刺はありませんが、南アジア~東南アジアに分布する祖先種は刺があります。これは人為選択によって刺が無くなったことを示しています。それだけではなく、ヒトの手によって形態が他にも大きく変わっていることが分かっています。これはヒトにとって便利になるように進化させたとも言えますし、ナスはヒトに保護してもらうように「労働寄生」しているともいう事ができ、進化の裏表の関係を示しています。こうして生まれたナスは最初はアジアと中東を中心に薬として用いられましたが、品種改良が進み、毒性が少なくなってからはヨーロッパ・アフリカ・アメリカも含め、未熟果を盛んに料理に使用されるようになりました。日本には飛鳥時代から奈良時代の間に渡来したようです。一方、ワルナスビは北アメリカ原産で勝手に生え、繁殖力が強く、刺と毒が多いことから対照的に嫌われる存在となっています。ナスとワルナスビは花についてはよく類似しており、特殊な雄しべの葯は特別なハナバチが振動することによってのみ、花粉を落とすことが分かっています。これは受粉の確実性を上げる工夫です。ワルナスビの果実は熟しても毒性を持ち続けており、これは自然界でもなぜ進化するか一見不思議な現象です。様々な説がありますが、研究によると、果実の栄養価が低いことと関係しているようです。本記事ではナスとワルナスビの分類・歴史・進化・繁殖生態・送粉生態・種子散布について解説していきます。
「ナス」の名がつく名前と花の形が類似した2種
ナス(茄子) Solanum melongena は別名ナスビ。諸説ありますがインド東部が原産とされる多年草で、日本では一年草として栽培されます。東南アジアでは古くから栽培されていました(講談社,2013)。近東とアジアで人気が出て、日本では奈良時代には中国から伝来しています(青葉,2013)。紫色の未熟果は主に食用とし(講談社,2013)、果実・茎・へた・根・花などは薬用にされることもありました(田中,1995)。
ワルナスビ(悪茄子) Solanum carolinense は別名オニナスビ・ノハラナスビ。北アメリカ原産の帰化植物で世界各地に帰化し、畑地・市街地・荒地・農耕地に生える多年草です。強害草として知られ、日本には初記録は1906年(明治時代)で、昭和初期から広がりました。和名は繁茂し駆除が難しく、果実にも毒性があることに由来します。
どちらもナス科ナス属で、名前や花の形が類似しているので、見たことがない人は2種を混同するかもしれません。
ナスとワルナスビの違いは?
しかし、ナスとワルナスビは全く異なる種類です。
ナスは栽培植物で自生することはありませんが、ワルナスビは野草として自生します。
形態についてはナスでは茎や葉には刺がなく、星状毛もないのに対して、ワルナスビでは茎や葉にはふつう刺があり、星状毛があります。
花に関してもナスでは花冠が紫色であるのに対して、ワルナスビでは花冠が白色~淡紫色です。
果実に関してもナスでは紫色から黄褐色に熟し縦長に肥大化し、熟すとソラニンが少量になり可食であるのに対して、ワルナスビでは緑色から橙黄色に熟し小型の球形で、熟してもソラニンを多量に含み食べられません。
原産国もナスはインドですが、ワルナスビは北アメリカですので、ナス属であること以外、殆ど共通点がないと言えるほど大きく違っています。








他に似た種類はいる?
ナス属はこの他にも多数種類が含まれていますが、草本で直立するナス属の仲間は限られています。
つる性や木本のナス属の種類を知りたい人は別記事を御覧ください。



ナスはどのように進化した?ナスも元々刺を持っていた!?
ナスはどのように進化したのでしょうか?
昔から様々な考察がなされてきましたが、現在ではこのような生き物の進化の過程を調べるために、遺伝子を用いて行う「分子系統解析」という科学的な手法が発達し、従来よりも正確に生き物同士の種類の近さを計ることができるようになっています。
ナスについても同様の手法を用いて調査が行われた結果、Solanum insanum という種類がナスに最も近いことが分かっています(Ranil et al., 2017; Aubriot et al., 2018)。そのため、この種類を見ると、ナスがどのように進化してきたのか分かってきそうです。
Solanum insanum は最近明確に分離された種類なので、完全に分布が明らかになっている訳ではありませんが、現在のところ、南アジア(パキスタン・インド・スリランカ)・東南アジア(タイ・ラオス・カンボジア・ミャンマー・インドネシア・フィリピン)・中国・台湾・マダガスカルで発見されています。
このことから南アジア~東南アジアがナスの原産であることは間違いないでしょう。ただし、マダガスカルのような隔離された分布も見られます。これは栽培のために人為的に導入されたものであると考えられています。
ナスと Solanum insanum の形態的な違いはなんでしょうか?
Solanum insanum は全草に刺を持っており、果実は球形で小さく、中果皮は海綿状ではありません。これに対して、ナスは上述のように刺はなく、果実は肥大化し、中果皮は海綿状です。
これはつまり、ナスは刺がなく、果実が大きく、海綿状の中果皮を持つように Solanum insanum から人為選択されたということを意味しているのです。

ナスに刺はあると、当然ヒトが育てる時に怪我をしてしまうかもしれませんし、果実や中果皮は発達したほうがヒトにとっては食べごたえがあるでしょう。
更に Solanum insanum とは違い、現在のナスは1つの花柄(花と茎を繋ぐ部分)に複数の花が咲いても、基部の1個以外は結実しないという特徴があります(田中,1995)。つまり、体の限られた資源を1つの果実に集約するようにも進化しているのです。
また、ソラニンの減少も後の時代の品種改良の時に起こったと考えられます。
これらはヒトによっては良いことばかりにも思えます。ところが、刺がなく、ソラニンが減少したことは明らかに外敵から身を守りにくくもなっています。実際、ナスが害虫や鳥に食べられやすいのは育てたことがある人からするととても共感できると思います。
これは言い換えると、「ナスはヒトによって外敵から守られながら育つことを前提に、防御を捨てて果実を作ることに特化して進化した」ということです。このような進化は栽培植物では広く知られています。
確かに現在のナスは刺を持っておらず、育てやすい一方で、常に目を配る必要があります。これは古代人の努力の成果とも言えますし、ナスがヒトに「労働寄生」しているということでもあるのです。ナスを育てる時はこのようなことを思い出すと進化の不思議を感じられそうです。
ナスの歴史は?元々は食用には適していなかった!?
ナスは2016年の総生産量が約5,100万トンで、トマトに次いで2番目に経済的に重要なナス科の果物作物です(Aubriot et al., 2018)。そんなナスはどのように世界に広まっていったのでしょうか?
上述のように Solanum insanum からナスが人為選択され、栽培化された訳ですが、その栽培化は複数地点で起こった可能性があります(Ranil et al., 2017; Aubriot et al., 2018)。その地域は東南アジアと南アジアが考えられます。一般的にはインド・ビルマ地域が栽培化の中心地であったと考えられています(Daunay & Janick, 2007)。興味深いことに現在日本人が食べる「水なす」とは違い、品種改良が進む前のナスはソラニンなどの毒性が強く薬用としての役割が強く、食用するにしても何らかの加工が必要だったようです。
インドでは紀元前300年頃(マウリヤ朝)のサンスクリット語の文献に、ナスに関連する様々な言葉が記されており、食用や薬として広く普及していたことが伺えます(Daunay & Janick, 2007)。サンスクリット語のインド・スリランカ発祥の伝統医療であるアーユルヴェーダでは、白い果実は糖尿病患者に、根は喘息の治療への使用が推奨されています。一方で、4世紀(グプタ朝)の古代ヒンドゥー教の経典『マールカンデーヤ゠プラーナ』では、ナスは好ましくないものの中に含まれています。現在のインド料理としては茄子のアチャールやベグニなどが知られます。
中国では早くから野菜作物として利用され、西晋時代(265〜316年)に嵇含 が著した最古の植物誌である『南方草木状』や南北朝時代(420〜581年)に北魏で6世紀頃に賈思勰によって書かれた農業書『斉民要術』でその存在が示されています。李時珍は1578年に成立した中国本草学の集大成である『本草綱目』内で果実・花托・根・茎・葉を用いた薬物製剤が膿瘍・腸管出血・歯痛など様々な病気を治すと述べています。現在の中国料理としては魚香茄子などが知られます。
日本へは8世紀頃、遣唐使が盛んになり、唐との関わりが深くなった時に伝わったと考えられています。文献上の最古の記録はよく『東大寺正倉院文書』(750年)であるとされていますが、『長屋王家木簡』(710年頃)には既に登場しています(森下,1999)。710年はちょうど藤原京から平城京に遷都し、飛鳥時代が終わり、奈良時代が始まった時です。『長屋王家木簡』では糠漬けでの利用が記されており、927年(平安時代中期)に成立した律令の施行細則をまとめた法典である『延喜式』からは生食や醤漬け・糠漬け・干し茄子として食されたことが分かります。最初は高位の者への進物として用いられていましたが、江戸時代頃より広く栽培されるようになり、ようやく庶民にもなじみのある野菜になりました。現在の日本料理としてはしぎ焼き・浅漬け・麻婆茄子・なす田楽・焼きなす・なすの味噌炒め・煮びたしなどが知られます。
ユーラシア大陸西方へは非常に早くペルシア(現在のイラン)に伝わっていますが、その時期は不明です。イスラーム世界が生み出した最高の知識人とも称されるサーマーン朝の学者イブン゠スィーナーが1020年に完成させた古来のアラビア医学にギリシアやインドの医学知識を加えた大著『医学典範』内でナスが登場しています。以降中世ペルシアを代表する医学・植物学の作家たちによって言及され、塩漬けによって毒性を抑え、果実を胆汁中和や耳病治療に用いることができるとも述べています。
ヨーロッパへの伝来は遅く、長らく古代ギリシアや古代ローマでは知られていませんでしたが、7〜8世紀のムスリムの進出により、地中海沿岸に広がり、ヨーロッパにまで広がりました。しかし、15世紀までは人気が出ませんでした(Santich & Bryant, 2008=2010)。これは毒性によるものも考えられますが、元々熱帯植物で栽培が難しかったことも理由の1つのようです(講談社,2013)。現在の地中海料理としては日本でも有名なフランス料理のラタトゥーユの他、ナスジャム(トルコ)・ババガヌーシュ(レバント)・アジャプサンダリ(ジョージアおよびアルメニア)・ムサカ(ギリシャ)・カポナータ(イタリア)・トゥンベット(スペイン)・ピスト゠マンチェゴ(スペイン)などがあります。
8世紀以降になると、ペルシアやアラブの船員によって東アフリカにも伝わっています。大航海時代(15〜17世紀)にスペインからアメリカへ野菜の1つとして運ばれ、17世紀半ばにはブラジルでも報告されています。
ナスの健康効果は?
ナス果実の93%以上は水分で、他の野菜と比べると低カロリーであるため、この点が評価を下げる点でもありますが、逆に言えば、食物繊維もあり、ダイエットには最適であると言えます。
ビタミンはほとんど含まれていないと言われることもありますが、ビタミンB2・ビタミンCや、カリウム・カルシウムなどのミネラル類は比較的少ないながらも、まんべんなく含まれています。
ナスの健康効果を網羅した研究では以下のような事例が挙げられています(Fraikue, 2016; Gürbüz et al., 2018)。ナスを量的にどのくらい摂取すれば良いのか?という点や他の食べ物に比べてどのように優れているのか?という点はまだ不明な点は多いですが、過剰摂取を避ければ大事な栄養源として体を助けてくれるでしょう。
- 鉄キレート剤として、妊婦・授乳中の母親・10代の特に女性に強く推奨される食材とされています。ナスに含まれる鉄分は月経前症候群・無月経・妊産婦貧血を改善することができます。
- 食物繊維は有害な化学物質や毒素を吸収する性質があるため、がん細胞の増殖や発達を抑制します。
- ナスニン(アントシアニン)という成分が体内に蓄積された鉄分を除去する作用があるため、鉄分を正常化し、コントロールする働きがあります。また、ナスニンは過酸化脂質から人間を守る働きがあります。
- 有害なフリーラジカルを除去し、心臓内の血管をほぐし、最終的に血液循環を強化することにより、心血管疾患(脳卒中・心停止・心臓病)の減少を助けます。食物繊維・カリウム・ビタミンB6・フラボノイドを豊富に含むナスは上記のような病気のリスクを低減させます。
- 摂取することでコレステロールを低下させる作用があります。クロロゲン酸は主要な抗酸化物質として作用し、豊富な食物繊維は肝臓で胆汁を生成するために血中コレステロールの吸収を促します。
- カリウムは電解質バランスを維持し、血圧のコントロールに必要なナトリウムの中和作用を助けるため、摂取すると血圧のコントロールが可能になります。
- 食物繊維がお腹にたまる食べ物のかさを作り、長い間満足感を保つので、ダイエットに用いることが出来ます。
- 髪や爪、肌を健康的に保ちます。含まれている高い水分・ビタミン・ミネラル・脂肪は、髪・爪・皮膚の質を高め、皮膚がんへの抵抗を含む身体の一般的な機能を向上させます。
- 高いα-グリコシダーゼとアンジオテンシンが体内のグルコースを吸収し、他の栄養素とともに2型糖尿病を制御します。また、フェノール豊富な抽出物は、糖尿病の制御に必要な酵素とグルコース同化を助けます。さらに、低炭水化物であり、食物繊維が血糖値のコントロールに役立ちます。
- 大量に摂取すると脳細胞の損傷を防ぐことができます。また、記憶機能を高め、細胞膜を保護する植物性栄養素も含まれています。
- マグネシウム・マンガン・カリウム・銅など、人間が必要とするミネラルを含んでいるため、骨の健康を促進します。
- 食物繊維が老廃物や有害な毒素を押し流し、健康的な消化を促進します。
- 脳細胞を保護し、フリーラジカル細胞の破壊から脳細胞を守り、脳腫瘍を回避する抗酸化物質がある一方で、心の健康を保ちます。含まれてるクロロゲン酸とフェノール物質は抗ウイルス・抗変異原性・抗菌物質として働き、健康な生活をコントロールするために、穏やかな抗体の供給を助けます。
- 含まれる食物繊維は腸の動きを活発にし、有害な毒素を掃除して、便秘を予防します。
ワルナスビはなぜ嫌われる?
ナスはこれだけヒトに利用されてきましたが、対照的なのはワルナスビです。名前の通り、悪者扱いされています。それはなぜなのでしょうか?
それは繁殖力が強く世界中の様々な環境に広まっている上に、駆除が難しく、刺があり、更に果実を含む全草に毒性があることにあります(宮崎ら,2011)。
原産地はアメリカ合衆国とカナダですが、ハイチ・ブラジル・バングラデシュ・グルジア・インド・ネパール・韓国・日本・クロアチア・ノルウェー・ニュージーランドにまで侵入しています。
日本におけるワルナスビの初記録は1906年(明治時代)の千葉県成田市の牧場ですが、海外から輸入した牧草種子に混入していたものとされています。しかし、この個体群がそのまま全国に広がったわけではないようです。
第二次世界大戦後(1945年~)になると、各地でワルナスビが記録されており、進駐軍の運搬した軍事物資に混入していた可能性が指摘されています。1990年代には飼料畑の強害雑草として広く知られています。これはアメリカ合衆国のトウモロコシ・ダイズ・ワタの強害雑草であることから、輸入濃厚飼料(たんぱく質・炭水化物・脂肪などの栄養素を多く含み、繊維質が少ない餌料)に由来する可能性があります。現在では北海道から沖縄まで分布は広がっています。ただし沖縄の記録は別種である可能性も指摘されています。
生息環境も畑地・草地・果樹園・畦・路傍・空き地・林縁・道路の植え込み・河川敷など人為的撹乱が入った所には極めて広く生えます。
そして繁殖力が強いです。ワルナスビは垂直および水平に広がる地下茎を張って、根系や茎の地下基部から新たなシュートを出す栄養繁殖を行います。そのため耕起作業によって地中の根から発芽したり、造成時の土砂や植木についた土等に根断片混入することで広がることができます。実際、京都市での研究では1km以上離れた地点でクローンが確認されています。全国への分布拡大は主にこのような経路を辿っているのかもしません。このような特徴は駆除を難しくしています。
また日本では稀な可能性がありますが、後述のように果実を生産し、種子散布による種子繁殖も行います。これは別株と一定の遺伝子の交換行うことで、新たな環境に適応するために行う必要があるのでしょう。
ここまでだけでも、景観などに影響を与える可能性がありますが、問題はそれだけではありません。全草に刺があるので、道を歩く時や、土地を利用するために人力で抜き取りや刈り取りを行う時に怪我をする可能性があります。このことも駆除を難しくしている要因です。更に収穫機へ絡みつくことや、家畜の草原の利用スペースを減らすといった問題も挙げられます。
また毒性があることも問題です。ナス科に共通のソラニン(solanin)、サポニン(saponin)などの毒素が含まれています。ソラニンが加水分解して生成するソラニジンは神経毒で、エストラーゼ阻害作用を持ち、家畜が採食すると、中毒症状として運動障害・痙攣・昏睡・黄疸などの肝臓障害を引き起こすことがあります。
同じナス属でもここまで評価が異なるというのは生物の進化の興味深い点です。
ナスとワルナスビの花の構造は?
ナスは非常に身近な植物ですが、その花は育てていないと見る機会が少ないかもしれません。花期は6~9月で、花序は殆どが単生花、まれに縮小した総状花序となっています。花序柄はほとんどありません。長さ1~1.8cmの花柄に、花冠が紫色で、長さ3~5cmの花を下向きに1個から数個咲かせます(田中,1995)。萼には星状の綿毛があり、しばしば、外面に長さ約3mmの刺を持ちます。
ワルナスビは花期が6〜10月。茎の途中から太い枝を出し、先端に直径約2cmの淡紫色または白色の花を6〜10個集散状につけます。花冠は浅く5裂して皿状に開きます。
ナスは上述のように1つの果実に栄養を集約するために、ワルナスビよりも花が少なくなっていますが、これを除けば2種含め、ナス属の花はかなり共通点が多い構造となっています。

ナス属の特徴的な葯はハチの振動に依存していた!?
特に雄しべにはナス属共通の大きな特徴があります。
それは雄しべの花粉を含む部分である葯が黄色で大きく目立ってることで、この葯は先端が細まり小さな穴が空いています。このような葯は「孔開葯」と呼ばれています(清水,2001)。このような特殊な葯はどのような役割があるのでしょうか?
これはナスを含むナス属の多くが「振動送粉」という特別な送粉を行うことと関係しています(Raju & Rao, 2006)。マルハナバチやクマバチなど一部のハナバチは大きな黄色い葯に止まった後、翅の筋肉で振動し、先端の穴からぽろぽろと花粉を落として運んでいきます。こうする事で、ナス属側はクマバチ属などのハナバチ類による盗蜜(萼などに穴を開けて花粉をつけずに直接蜜だけを奪う行為)を防ぎ、彼らに花粉だけを与えて送粉させることができます(市川ら,2011)。またあちこちの別種の植物の花に訪れ、受粉を不確実にするハナアブ科やミツバチのような昆虫が訪れるのを制限することが出来ます。
下向きに咲くこともやってくる昆虫をハナバチに限定させていると考えられます。下向きの花にうまくぶら下がるには脚力と飛翔力が必要で、そのような能力を持っているのは一部のハナバチだけです。
更にナス属は蜜を作らないので、ハナバチに花粉を与えることに特化し、チョウや蛾などが訪れないように進化してると考えられます。
このようにナス属はやってくる昆虫を限定することで巧みに受粉を確実性を上げていたのです。
具体的にはワルナスビについては日本ではクロマルハナバチ Bombus ignitus がやってきた記録があります(市川ら,2011)。
原産地のアメリカ合衆国ペンシルベニア州でも花に訪れた昆虫146匹のうち138匹がマルハナバチ属の一種 Bombus impatiens でした(Kariyat et al., 2021)。ただし、コネチカット州ではコハナバチ科のコハナバチ属の複数種 Lassioglosum spp. や Augochloropsis metallica も訪れるとされています(Quesada-Aguilar et al., 2008)。
なぜ同じ個体内に雌しべが長いタイプと短いタイプがあるのか?
ナスの花は同一個体の中で雌しべが雌しべより長いタイプ、同じ長さのタイプ、短いタイプの3タイプが存在することが知られています(農業・生物系特定産業技術研究機構,2006)。
この長さは雌しべの中でも花柱の部分の長さが違うので、雌しべが雌しべより長いタイプの花を「長花柱花」、雌しべが雄しべと同じ長さのタイプの花を「中花柱花」、雌しべが雌しべより短いタイプの花を「短花柱花」と呼ぶことがあります。
また、ワルナスビについても長花柱花と短花柱花の2タイプが存在することが明らかになっています(Quesada-Aguilar et al., 2008)。
このような花柱の長さが違う性質を「異型花柱性(heterostyly)」と呼び、様々な植物で見られます(渡邊,2022)。

しかし、同じ異型花柱性でもナスやワルナスビは他の植物とは異なる点があります。それは、他の植物では「別個体で」異型花柱性が起こっているので、普通個体内ではどちらかの花柱の長さのタイプしか見られません。それに対して、ナスやワルナスビでは「個体内で」異型花柱性が起こっているのです。
なお、このような状態を雄性両性同株(andromonoecy)ということもありますが、あまり適切とは言えません。
「別個体で」起こる異型花柱性は一般に自家受粉を防ぐ手段であると考えられています。
では「個体内で」起こる異型花柱性はどのような役割があるのでしょうか?
いくつか仮説があるのですが、ワルナスビを対象にしたアメリカ合衆国コネチカット州で行われた研究によると、このように花柱の長さを変えることで、それぞれの花で雄としての役割と雌としての役割を分担しているという考えが支持されています。
つまり、長花柱花では雌しべが雄しべより長いため、雌しべが突き出している状態になります。そうなると雌しべが邪魔になり、うまくハナバチが振動受粉することが出来ず、雄しべから花粉を取ることが出来ませんが、ハナバチが花にしがみついたときに別の個体の花粉を受け取ることができます。したがって長花柱花は雌としての役割を担い、果実を作ることになります。
一方、短花柱花では雌しべが雄しべより短いため、雌しべは雄しべの中に埋もれてしまい、受粉することはできません。その代わり、ハナバチは邪魔なものがないので効率的に花を振動させ花粉を落として体に付けていきます。したがって短花柱花は雄としての役割を担い、花粉を他の個体に広げることに専念しているのです。
興味深いことにこの作用はマルハナバチ属の一種 Bombus impatiens が訪れた時にのみ発揮されることも分かっています。
コネチカット州ではコハナバチ科の仲間もワルナスビの花に訪れることが分かっていますが、 Bombus impatiens の場合はコハナバチ科の仲間よりも一回の振動の時間も短く、何度も花に触れ、効率的に花粉を運びます。そのため、長花柱花と短花柱花での花粉分散効率の違いが数字として顕著に出たと考えられます。
ナスやワルナスビの花は3タイプあると言いましたが、必ずしも全てあるわけではなく、1つのタイプしか付けていない場合もあり、個体差が激しいことも分かっています。
これはつまりやってくるハナバチによって異型花柱性を発生させた方が良い場合とさせない方が良い場合があるということを示していると考えられます。マルハナバチがやってくる環境であるということが、「個体内で」起こる異型花柱性の発生と密接にリンクしている可能性があるのです。
このような訪花昆虫が植物に与える影響の研究が進めば、なぜ異型花柱性を持つ植物と持たない植物が居るのかもっと一般的なことが分かってくるでしょう。
ナスとワルナスビの果実の構造は?
ナス属の果実は共通で液果です。
ナスも液果に分類されますが、同じナス属のトマトとは違い甘味のあるものではありません。黒色~紫色で完全に熟して黄褐色、形や大きさは品種によって違い、非常に変化が多いですが、ほとんど直径6cm以上です。種子はレンズ形、黄色を帯びます。
ワルナスビの液果は直径約1.5cmの球形で緑色から黄色に熟します。
ナスの果実は食用に大きく発達したわけですが、どのような構造になっているのでしょうか?
ナスの果実は外果皮・内果皮・隔壁(partition)・胎座(placenta)・果心(core)の入れ子構造になっています(Wang et al., 2021; 中村,2021)。
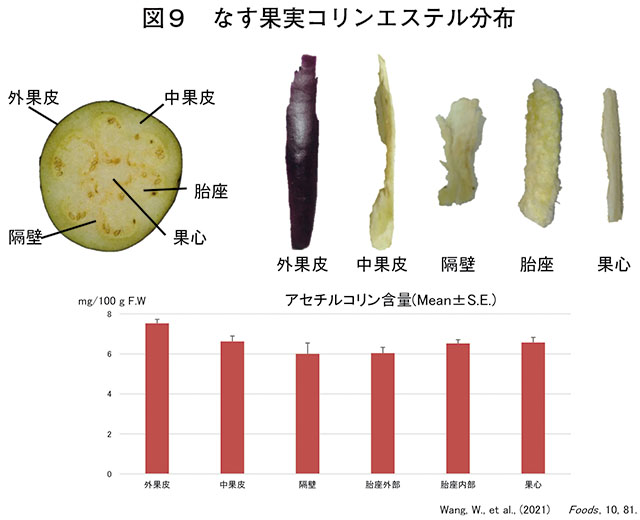
食用にする未熟果の一番外側の紫色の部分が「外果皮」にあたります。外果皮には約4種類のアントシアニンが含まれており、その含有量の90%がモノアシルアントシアニンであるナスニン(nasunin)で、紫色を作り出す主成分です(伊藤ら,2013)。
その内側部分は全て白く、海綿状なため、目で正確に判別するのは難しくなっています。
外果皮の内側には肉厚な内果皮があります。その内側は「隔壁」によって分離されてた「胎座」が複数あり、胎座には種子が多数置かれています。複数の胎座は果実の中心部分である「果心」を囲んでいます。
ワルナスビの種子散布方法は?なぜ果実に毒があるのに食べる動物がいる?
ナスの種子は現在では上述のようにヒトの手によって散布されていると考えてよいでしょう。
ワルナスビはどのように種子を散布するのでしょうか?
具体的な動物の種類まで記録した観察はありませんが、北アメリカの研究では鳥と哺乳類によって果実が食べられると考えられています(Cipollini & Levey, 1997a)。しかし日本ではワルナスビの果実をおいて赤外線カメラで観察した実験がありますが、食べる動物は確認されていません(宮崎ら,2011)。ただ、私は車道沿いの植樹帯に孤立して生えるワルナスビを見たことがあります。これは移植した土にワルナスビの根が混入していた可能性もありますが、鳥によって果実が食べられている可能性もあります。
ところで、ワルナスビの果実には大きな疑問があります。上述のようにワルナスビの果実には熟してもソラニンを含み、毒性があります。これは全てのナス属の植物がそうなっているわけではなく、熟すとソラニンは無くなることも多いのです。なぜこのような熟してもまずい果実が実在するのでしょうか?
この不可解な現象については詳しく研究が行われています(Cipollini & Levey, 1997a; b)。
まず最初に考えられるのは、毒性によってやってくる動物を制限している可能性です。この果実はヒトが食べても毒性がありますし、家畜が摂取することで運動障害・痙攣・昏睡・黄疸が発生することも分かっています。一方で、種子を破壊しないように果肉のみを消化したり、代謝が異なる鳥や一部の野生哺乳類では無毒なのかもしれません。これならば特定の動物にのみ果実を食べさせて、種子散布させることが可能でしょう。このような仮説を「直接毒性仮説(Directed Toxicity Hypothesis)」と言います。
このことを確かめるために、北米で一般的な果実食を行う鳥であるコマツグミ Turdus migratorius、コリンウズラ Colinus virginianus、 哺乳類であるシカシロアシネズミ Peromyscus maniculatus、キタオポッサム Didelphis virginiana の4種にワルナスビの果実とともに、無毒なテリミノイヌホオズキ Solanum americanum の果実を実際に与え、摂食率が比較されました。
その結果、いずれの哺乳類や鳥類であってもテリミノイヌホオズキよりも有毒なワルナスビの果実を嫌い、摂食率は大きく低下していたのです。これはつまり、直接毒性仮説のように、特定の動物のみに無毒であるという可能性が低いことを示しています。
だとすると他にはどのような理由が考えられるでしょうか?これには更に2つの仮説が存在し、それは果実の栄養価と関係を考慮したものです。
まず1つ目の仮説としては、「除去率仮説(Removal Rate Hypothesis)」という考えです。もし生息環境に鳥や哺乳類が少なかったり、相対的に他の植物より果実の栄養素が少なく人気がない場合、いつまでも果実は植物体についたままになります。その場合、害虫や真菌によって果実が侵されてしまう可能性が高くなるでしょう。そうであるならば、毒性を持ってこれらの害のある生き物を退けるメリットが、種子散布に貢献する動物に食べられないというデメリットを上回ることになります。
逆に生息環境に鳥や哺乳類が多かったり、相対的に他の植物より果実の栄養素が多く人気な場合、さっさと果実は鳥や哺乳類に食べられ、害虫や真菌から身を守る必要はありません。この場合、毒性を減らすと、種子散布に貢献する動物に食べられるというメリットが害のある害虫や真菌を退けられないデメリットを上回ることになります。
端的に言うと、果実が食べられる率が低いなら毒性が高くなり、食べられる率が高いなら毒性が低くなるということです。
2つ目の仮説としては、「栄養価-毒素滴定仮説(Nutrient-Toxin Titration Hypothesis)」という考えです。こちらは対照的な考えです。もし栄養価が高く、鳥や哺乳類に人気がある場合、その分毒性を高めることで害虫や真菌からの防御にも投資できるかもしません。一方、栄養価が低く、鳥や哺乳類に人気がない場合、毒性も低くしなければ、鳥や哺乳類には見向きもされない可能性があります。
こちらの仮説は端的に言うと、果実が食べられる率が高いなら毒性が高くなり、食べられる率が低いなら、毒性が低くなるということです。
どちらも説得力があるように見えますが、これは実際に実験してみることで確かめられそうです。
直接毒性仮説を検証した同じ研究で、このことも実験されています。その結果、毒性が強いワルナスビの果実が栄養価が高いという事実は認められませんでした。このことは「除去率仮説」を支持しています。
まとめると、ワルナスビの果実は生息環境に鳥や哺乳類が少なかったり、相対的に他の植物より果実の栄養素が少なく人気がないなどの理由により、長期間害虫や真菌から防衛する必要性が高まり、例え鳥類や哺乳類に嫌われたとしても毒性を高めていたのです。
ワルナスビは上述のように荒地に生えることに特化しているので、果実の栄養価を十分高めるのは難しかったのかもしれません。そのため苦肉の策として果実をソラニンなどのグリコアルカロイドで守っているのでしょう。毒があることで悪者扱いされることもありますが、このようなワルナスビ側の事情があることも知るとまた違った姿が見えてくると思います。
引用文献
Aubriot, X., Knapp, S., Syfert, M. M., Poczai, P., & Buerki, S. 2018. Shedding new light on the origin and spread of the brinjal eggplant (Solanum melongena L.) and its wild relatives. American Journal of Botany 105(7): 1175-1187. https://doi.org/10.1002/ajb2.1133
青葉高. 2013. 日本の野菜文化史事典. 八坂書房, 東京. 486pp. ISBN: 9784896941609
Cipollini, M. L., & Levey, D. J. 1997a. Why are some fruits toxic? Glycoalkaloids in solanumand fruit choice by vertebrates. Ecology 78(3): 782-798. https://doi.org/10.1890/0012-9658(1997)078[0782:WASFTG]2.0.CO;2
Cipollini, M. L., & Levey, D. J. 1997b. Antifungal activity of Solanum fruit glycoalkaloids: implications for frugivory and seed dispersal. Ecology 78(3): 799-809. https://doi.org/10.1890/0012-9658(1997)078[0799:AAOSFG]2.0.CO;2
Daunay, M. C., & Janick, J. 2007. History and iconography of eggplant. Chronica Horticulturae 47(3): 16-22. ISSN: 0578-039X, https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=79218125888314fab4f5275db48d5625b124c915
Fraikue, F. B. 2016. Unveiling the potential utility of eggplant: A review. Conference Proceedings of INCEDI 1: 883-895. https://www.researchgate.net/publication/310846026
Gürbüz, N., Uluişik, S., Frary, A., Frary, A., & Doğanlar, S. 2018. Health benefits and bioactive compounds of eggplant. Food Chemistry 268: 602-610. https://doi.org/10.1016/j.foodchem.2018.06.093
市川俊英・倉橋伴知・幾留秀一. 2011. 香川県で採集された訪花ハナバチ類とキムネクマバチによる新奇な授粉様式の可能性. 香川大学農学部学術報告 63(116): 43-59. ISSN: 0368-5128, http://id.nii.ac.jp/1731/00003553
伊藤和子・阿久津智美・大山高裕・渡邊恒夫・山﨑公位・角張文紀・宇田靖. 2013. ナス下漬液からナスニンおよびクロロゲン酸を含む抗酸化性粉末の調製. 日本食品科学工学会誌 60(1): 30-37. https://doi.org/10.3136/nskkk.60.30
Kariyat, R. R., Bentley, T. G., Nihranz, C. T., Stephenson, A. G., De Moraes, C. M., & Mescher, M. C. 2021. Inbreeding in Solanum carolinense alters floral attractants and rewards and adversely affects pollinator visitation. American Journal of Botany 108(1): 74-82. https://doi.org/10.1002/ajb2.1594
講談社. 2013. 野菜の本 からだにやさしい旬の食材. 講談社, 東京. 255pp. ISBN: 9784062183420
宮崎桂・西田智子・浦川修司. 2011. 雑草モノグラフ 6. ワルナスビ (Solanum carolinense L.). 雑草研究 56(3): 154-165. https://doi.org/10.3719/weed.56.154
森下正博. 1999. ‘水茄子’の果実特性と来歴. 大阪府立農林技術センター研究報告 35: 34-39. https://agriknowledge.affrc.go.jp/RN/2010691355
中村浩蔵. 2021. なすの食品機能と機能性表示食品. 野菜情報 211: 40-48. ISSN: 2436-9586, https://vegetable.alic.go.jp/yasaijoho/senmon/2110_chosa1.html
農業・生物系特定産業技術研究機構. 2006. 最新農業技術事典. 農山漁村文化協会, 東京. v, 2003pp. ISBN: 9784540051630, https://lib.ruralnet.or.jp/nrpd/#koumoku=13429
Quesada-Aguilar, A., Kalisz, S., & Ashman, T. L. 2008. Flower morphology and pollinator dynamics in Solanum carolinense (Solanaceae): implications for the evolution of andromonoecy. American Journal of Botany 95(8): 974-984. https://doi.org/10.3732/ajb.0800106
Raju, A. S., & Rao, S. P. 2006. Nesting habits, floral resources and foraging ecology of large carpenter bees (Xylocopa latipes and Xylocopa pubescens) in India. Current Science 90(9): 1210-1217. ISSN: 0011-3891, https://www.jstor.org/stable/24092022
Ranil, R. H., Prohens, J., Aubriot, X., Niran, H. M. L., Plazas, M., Fonseka, R. M., … & Knapp, S. 2017. Solanum insanum L.(subgenus Leptostemonum Bitter, Solanaceae), the neglected wild progenitor of eggplant (S. melongena L.): a review of taxonomy, characteristics and uses aimed at its enhancement for improved eggplant breeding. Genetic Resources and Crop Evolution 64: 1707-1722. https://doi.org/10.1007/s10722-016-0467-z
Santich, B. & Bryant, G. 2008. Edible: an illustrated guide to the world’s food plants. National Geographic, Washington, D.C. 360pp. ISBN: 9781426203725 [= 2010. 世界の食用植物文化図鑑 起源・歴史・分布・栽培・料理. 柊風舎, 東京. 360pp. ISBN: 9784903530352]
清水建美. 2001. 図説植物用語事典. 八坂書房, 東京. xii, 323pp. ISBN: 9784896944792
田中孝治. 1995. 効きめと使い方がひと目でわかる 薬草健康法. 講談社, 東京. 123pp. ISBN: 9784061953727
Wang, W., Yamaguchi, S., Suzuki, A., Wagu, N., Koyama, M., Takahashi, A., … & Nakamura, K. 2021. Investigation of the distribution and content of acetylcholine, a novel functional compound in eggplant. Foods 10(1): 81. https://doi.org/10.3390/foods10010081
渡邊謙太. 2022. 「異型花柱性」を巡る生態学と進化生物学の今. 沖縄工業高等専門学校紀要 16: 31-45. https://doi.org/10.51104/nitokinawacollege.16.0_31
出典元
本記事は以下書籍に収録されたものを大幅に加筆したものです。

