
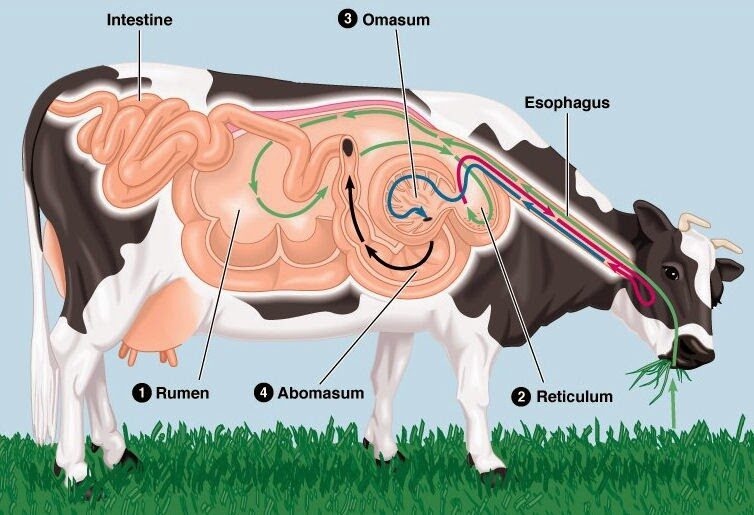
When you go to a yakiniku (Japanese barbecue) restaurant, you may come across four parts of the cow's stomach (fore-stomach) as a type of offal: mino, hachinosu, senmai, and giara.
Because it has a distinctive taste and is a type of offal, it's a love-it-or-hate-it kind of dish, but some people enjoy eating it because of its crunchy texture and how well it pairs with alcohol.
If you only eat them, you might not think much about their true nature or role, but they actually play an important role for ruminants, including cows.
This is essential knowledge for anyone involved in livestock farming, and it wouldn't hurt for the general public to know it as common sense.
However, the stomachs of ruminants have an overwhelming number of names, and I often find it difficult to understand the correspondence between their function and shape.
However, regardless of the sheer number of names, a closer examination reveals that the form, function, and order of these elements are highly logical and inevitable.
Here, I'll summarize some simple ways to remember the names and morphologies of the stomachs of ruminants.
To put it simply, the process is as follows: tripe (fermentation) → honeycomb tripe (separation) → omasum (absorption) → abomasum (digestion).
- What is rumination? Why do animals ruminate and have a forestomach?
- What are the different types of ruminants?
- List of names and functions of the stomachs (fore-stomachs) of ruminants
- How do you remember the name and function of the first stomach (rumen, tumor, or stomach tumor)?
- How can you remember the name and function of the second stomach (reticulum, honeycomb tripe, or stomach cavity)?
- How do you remember the name and function of the third stomach (lobe stomach, omason)?
- How can you remember the name of the fourth stomach (abomasum, giara, or abomasum) and what is its function?
- References
What is rumination? Why do animals ruminate and have a forestomach?
Rumination is the act of bringing grass that has been swallowed back into the mouth and chewing it again.
If you observe cows, you'll notice them constantly chewing for extended periods. This is because their stomachs are regurgitating food they've already swallowed, and they're chewing it again.
While this behavior may seem a little unpleasant to humans, it is an adaptation that gives herbivorous ruminants a significant advantage in grasslands.
The grass that is regurgitated does not simply return to the mouth as is. Ruminants have a stomach divided into four compartments, and the first stomach (the rumen) houses microorganisms, which ferment the grass they eat. Furthermore, the regurgitated grass is not exposed to stomach acid.
The plant cell walls containing polysaccharides such as cellulose and hemicellulose found in grass are extremely strong and normally only partially digestible by the digestive capabilities of ordinary animals. However, ruminants produce volatile fatty acids (VFAs, such as acetic acid, propionic acid, and butyric acid) through fermentation, which serve as a source of nutrition (Mackie, 2002; Bao et al., 2019).
However, microorganisms cannot properly ferment large pieces of grass. Therefore, ruminants repeatedly bring the grass back into their mouths, physically destroying it with their teeth and thus assisting microbial fermentation (Matsuda et al., 2011).
This allows them to efficiently obtain nutrients from even small amounts of grass, surpassing other mammals (Hofmann, 1989).
Furthermore, while microorganisms themselves synthesize proteins for their own use, ruminants send urea from their food and urea synthesized in their liver to their first stomach, allowing microorganisms to synthesize proteins, which the ruminants then swallow, making them their own proteins (Hailemariam et al., 2021).
Furthermore, it has been pointed out that stomach microorganisms can break down some plant secondary metabolites (phenols, alkaloids, etc.), which helps broaden their diet and increase their tolerance to toxicity (Loh et al., 2020).
While this may seem like a highly efficient function, it is a function completely specialized for herbivores. For frugivorous, carnivorous, or omnivorous animals, it lacks flexibility and is not cost-effective to maintain such a voluminous stomach (a trade-off). Therefore, humans and pigs do not have such stomachs.
What are the different types of ruminants?
Academically, "true ruminants" (Ruminantia), which have four stomachs (quadriventricular stomachs), refer to animals belonging to the suborder Ruminantia of the order Artiodactyla. The suborder Ruminantia is divided into two suborders, the infraorder Machodea and the infraorder Euruminantia, and six families, with approximately 200 extant species (Bao et al., 2019). The major families and representative examples are as follows:
- Bovidae: Includes genera such as cattle, sheep, goats, antelopes, and antelopes.
- Family Cervidae: Includes genera such as deer (Japanese deer), moose, and reindeer.
- Family Giraffidae: Giraffes and okapis.
- Pronghorn family (Antilocapridae): Pronghorn.
- Family Moschidae: Includes the genus *Musk deer*, etc.
- Family Tragulidae: Includes the genus Tragulidae, etc.
However, there are other animals that perform rumination (chewing the cud) and foregut fermentation, even though they do not have a four-chambered stomach. These animals have evolved convergently from different lineages and are therefore not usually considered "ruminants." Compared to ruminants, they have simpler stomachs and are less efficient at absorbing nutrients from grass.
The Camelidae family (camels, llamas, alpacas, etc.) belongs to the order Cetartiodactyla, but is part of the suborder Neopoda (cameloid suborder). They generally perform digestion that is "similar to rumination," but their stomach has three chambers and lacks the absorption function of the third stomach (lobular stomach) in ruminants (Fowler, 2008). They evolved separately from the ruminant suborder.
Although hippopotamuses belong to the order Cetartiodactyla, they are closely related to whales as they belong to the order Cetacea-Hippomorpha, and while they do not ruminate, they perform forestomach fermentation, which involves microbial fermentation in the forestomach (Clauss et al., 2004).
Many large marsupials (e.g., red kangaroos, large Australian macropods) perform foregum fermentation in the foregum, but do not engage in rumination in the usual sense (regular, repetitive re-chewing) (Vendl et al., 2017). However, there are individual records of regurgitation and re-chewing, which is called pseudo-rumination or merycism. Merycism is much shorter and less consistent than rumination, and is therefore often distinguished from rumination.
The proboscis monkey, Nasalis larvatus, also feeds on leaves and fruits and performs mericism and foregum fermentation (Matsuda et al., 2011).
The hoazin (Opisthocomus hoazin) is a bird, but it is a very unique example of a bird that performs strong microbial fermentation in an enlarged foregum (crop). Its chemical fermentation products (volatile fatty acids) and microbial communities are similar to those of mammals that ferment their foregums, but it does not ruminate (Grajal, 1995). It has been reported that it feeds its offspring substances that have been partially broken down and fermented from its food.
List of names and functions of the stomachs (fore-stomachs) of ruminants
Below is a table summarizing the naming of ruminant stomach parts based on their order, appearance, and their roles in relation to grilled meat.
It can be seen that fermentation, separation, absorption, and digestion occur from the first to the fourth stomach.
| Naming by order | Names based on appearance | Naming at a yakiniku restaurant | English name | role |
|---|---|---|---|---|
| rumen | Stomach tumor | Mino (straw raincoat) | Lumen | Compared to other parts of the stomach, it is noticeably enlarged in a lumpy shape, and microorganisms residing in the papillae ferment cellulose and starch, producing volatile fatty acids. |
| second stomach | Reticulum (stomach of the reticular cell) / Honeycomb stomach (stomach of the honeycomb cell) | Honeycomb | Reticulum | The mesh-like beams separate small particles and liquid phases and send them to the next passage, while sending larger particles back to promote rumination (re-chewing). |
| third stomach | Lobe stomach / Double-lipped stomach | Senmai (thousand slices) | Omasum | It absorbs water, electrolytes, and some volatile fatty acids from numerous leaves, increasing the solid content ratio of the digested food before transporting it to the fourth stomach. |
| abomasum | Wrinkled stomach / True stomach | Giara (false belly), Gyaras, Akasenmai, Abomi | Abomasum | The "true stomach," which has a glandular mucosa and is responsible for digestion, secretes hydrochloric acid and pepsin to hydrolyze microbial proteins and food proteins with acid. |
How do you remember the name and function of the first stomach (rumen, tumor, or stomach tumor)?

The first stomach is called a tumorous stomach in terms of its appearance, tripe in yakiniku (Japanese barbecue), and rumen in English.
The most distinctive feature of the first stomach is its significantly enlarged, lumpy appearance compared to the other stomach parts, which is why it is called a tumor stomach.
The name "mino" in yakiniku (Japanese barbecue) comes from the fact that when it's opened up, it resembles a "straw raincoat" (mino).
This morphology is closely related to the role of the first stomach.
The first stomach contains microorganisms (bacteria, protozoa, archaea, and fungi) that ferment cellulose and starch, producing volatile fatty acids (VFAs; acetic acid, propionic acid, and butyric acid). VFAs are the main energy source, and the microorganisms also supply proteins and vitamins (Perez et al., 2024). Lumen gas is expelled through movement (stirring) and belching (eructation).
In other words, it is thought that the first stomach is specially enlarged, like a "bump" or "skin," to expand the habitat for fermentation-promoting microorganisms.
Furthermore, numerous papillae develop on the inner surface of the rumen, increasing the surface area and aiding in VFA absorption (Pokhrel & Jiang, 2024).
The English word "rumen" is borrowed from the Latin word "rūmen." While there are several theories about its etymology, it is generally believed to originate from the Proto-Italic *roug(s)mən ~ *rug(s)mən < Proto-Indo-European *h₁rewg-, *h₁rewǵ- "to burp" (Wiktionary contributors, 2026).
This name also accurately describes the role of the rumen, where methane is mainly produced by the reduction of hydrogen (H₂) and methylated compounds by methanogenic archaea. This generates rumen gas, which mainly consists of carbon dioxide (CO₂) 45-75% and methane (CH₄) 20-35%, and ruminants expel this gas by belching (Zhang et al., 2020).
While burps containing methane from cows are sometimes considered a problem due to their role as greenhouse gases, this can be attributed to the rumen (first stomach).
The first stomach can be easily remembered if you think of it as a lumpy, cloven structure that houses microorganisms and is where unnecessary burps are produced.
As mentioned above, microorganisms also play a role in protein synthesis and detoxification of plant secondary metabolites (phenols, alkaloids, etc.).
How can you remember the name and function of the second stomach (reticulum, honeycomb tripe, or stomach cavity)?

The second stomach is called the reticulum in English, while the tripe is called honeycomb tripe in grilled meat.
The terms "reticulous stomach" and "honeycomb stomach" come from the fact that the mucous membrane of the stomach lining is arranged in a hexagonal, honeycomb-like pattern.
The English word "reticulum" is borrowed from the Latin word "rēticulum," meaning "net," and derives from the Latin word "rēte" ("net, snare") + -culum (diminutive suffix) < Proto-Italic *rēti- < Proto-Indo-European *h₁reh₁- meaning "separate, loose." It is cognate with the English word "reticle," which refers to the aiming line on a gun scope or microscope (Wiktionary contributors, 2026).
In other words, all of these originate from the fact that they have a polygonal, network-like structure.
This morphology is also deeply related to the role of the second stomach.
The second stomach contracts to separate small particles and the liquid phase and send them to the next passage, while sending back larger particles for rumination (re-chewing) (Song et al, 2024). The rumen and reticulum function in conjunction and are collectively called the "reticulorumen" or "ruminoreticulum."
During this sorting process, the multi-chambered spatial arrangement created by the pillars, which are sometimes called honeycomb or mesh-like structures, acts like a "shallow cage," controlling the flow rate and sorting efficiency.
In other words, larger particles remain inside the basket and are returned to the first stomach, while smaller particles and liquids flow to the third stomach and beyond. Conversely, digestive function is minimal.
Due to these characteristics, foreign objects such as metal can easily become lodged, which can lead to a disease called wound reticulogastroperitonitis (hardware disease), especially in cattle.
The second stomach is easier to remember if you think of it as an organ that sorts out food that needs to be ruminated, with a honeycomb-like network structure.
How do you remember the name and function of the third stomach (lobe stomach, omason)?

The third stomach is called "leaf stomach" in terms of its appearance, "senmai" in yakiniku (grilled meat), and "omason" in English.
The term "leaf stomach" comes from its structure, which consists of many layers of folds (thin, plate-like tissues) that resemble leaves or petals. The term "double-leaf stomach" is similar.
The name "senmai" is said to originate from its appearance, which looks like it has "a thousand" folds overlapping inside. There is also a theory that it comes from the Korean word "jeonyeob" (千葉/千枚), but the meaning is roughly the same.
In other words, they stem from almost the same characteristics.
This morphology is also deeply related to the role of the third stomach.
The third stomach absorbs water, electrolytes, and some VFAs, increasing the solid content ratio of the digested food before sending it to the fourth stomach. It can be said that it removes liquid and separates even finer particles (Pérez et al., 2023).
At this time, a large absorption surface is created by numerous leaves (laminae), enabling efficient absorption of water and VFAs.
The third stomach is easier to remember if you think of it as having countless leaf-like folds that appear to be thousands of leaves, and that it absorbs the liquid portion before the intestines.
The word omasum is a loanword from the Latin omāsum, meaning "bull's stomach." The Latin word is thought to be derived from Gaulish, which in turn was borrowed from Carthaginian, which in turn is thought to be derived from the Proto-Semitic *ḥamṯ- meaning "abdomen" (Wiktionary contributors, 2026).
The meaning of "Omason" has changed significantly from its original meaning.
How can you remember the name of the fourth stomach (abomasum, giara, or abomasum) and what is its function?
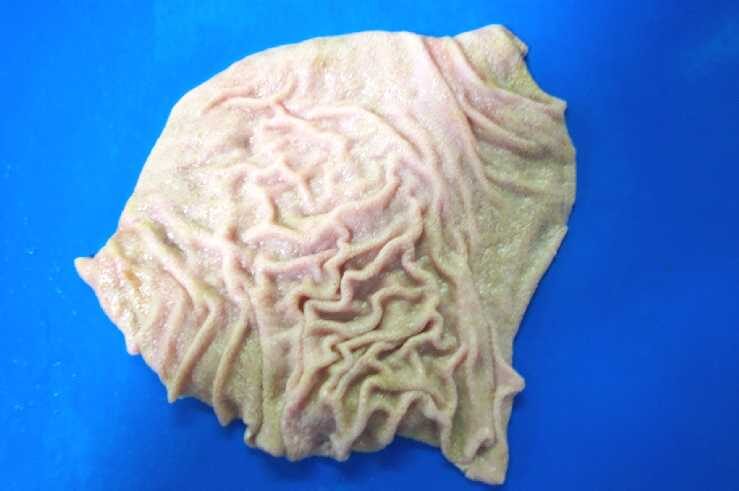
The fourth stomach is called "wrinkled stomach" based on its appearance, "giara" in the context of grilled meat, and "abomasum" in English.
The term "wrinkled stomach" comes from the fact that the mucous membrane has fine folds.
The term "true stomach" comes from the fact that it has the same function as the normal stomach of animals other than ruminants.
The fourth stomach is a "true stomach" with a glandular mucosa that secretes hydrochloric acid and pepsin, which acid-hydrolyze microbial and dietary proteins to prepare them for absorption in the small intestine (Ash, 1961). In this sense, it is "genuine."
Furthermore, the presence of wrinkles in the fourth stomach also has significance.
The folded folds in the stomach are folded when empty and unfold when full, increasing the gastric volume (variable volume). The folds also improve the efficiency of stomach wall movement (mixing and stirring), increasing the contact surface between acid and contents, thus facilitating digestion. These are not unique features and are consistent with the general function of the "rugae" of the stomach, including in humans.
Furthermore, the folding of the mucous membrane increases the surface area that accommodates the gastric glands, which secrete gastric juices such as hydrochloric acid and pepsin, allowing for more efficient secretion of acids, enzymes, and mucus.
Therefore, it's easier to remember that the fourth stomach is a "true stomach" with the same chemical digestion function for proteins as other mammals, and that it has gastric glands in its wrinkled folds that serve to temporarily store food.
Abomasum originates from the New Latin abomāsum < ab- "away from" + omāsum "bull's stomach." Therefore, it means "stomach away from the omason (third stomach)" (Wiktionary contributors, 2026).
As for the name "giara," a theory circulating on the internet suggests that it originated from "gibara," meaning "false stomach," because it functions like an intestine despite being the stomach.
However, it is the third stomach that functions like an intestine, while the fourth stomach is the stomach itself, as mentioned above, so this origin theory is not very convincing. It would be more understandable if it was confused with the third stomach, but there is no evidence to support that.
Another theory suggests that the name originated from "gyara" (gyaru) → "giara" (giara) because it was a payment (guarantee) received by people working at U.S. military bases (Tanabe, 2016), but the historical background is unclear, and this theory is also questionable.
References
Ash, RW 1961. Acid secretion by the abomasum and its relation to the flow of food material in the sheep. The Journal of Physiology 156(1): 93-111. https://doi.org/10.1113/jphysiol.1961.sp006660
Bao, W., Lei, C., & Wen, W. 2019. Genomic insights into ruminant evolution: from past to future prospects. Zoological Research 40(6): 476-487. https://doi.org/10.24272/j.issn.2095-8137.2019.061
Clauss, M., Schwarm, A., Ortmann, S., Alber, D., Flach, EJ, Kühne, R., … & Hofer, H. 2004. Intake, ingesta retention, particle size distribution and digestibility in the hippopotamidae. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 139(4): 449-459. https://doi.org/10.1016/j.cbpb.2004.10.002
Fowler, ME 2008. Camelids are not ruminants. In: ME Fowler, & RE Miller (Eds.), Zoo and Wild Animal Medicine (6th ed., pp. 375-385). Saunders. ISBN: 9781416057598, https://doi.org/10.1016/B978-141604047-7.50049-X
Grajal, A. 1995. Structure and function of the digestive tract of the hoatzin (Opisthocomus hoazin): a folivorous bird with foregut fermentation. The Auk 112(1): 20-28. https://doi.org/10.2307/4088763
Hailemariam, S., Zhao, S., He, Y., & Wang, J. 2021. Urea transport and hydrolysis in the rumen: a review. Animal Nutrition 7(4): 989-996. https://doi.org/10.1016/j.aninu.2021.07.002
Hofmann, RR 1989. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system. Oecologia 78(4): 443-457. https://doi.org/10.1007/BF00378733
Loh, ZH, Ouwerkerk, D., Klieve, AV, Hungerford, NL, & Fletcher, MT 2020. Toxin degradation by rumen microorganisms: a review. Toxins 12(10): 664. https://doi.org/10.3390/toxins12100664
Mackie, RI 2002. Mutualistic fermentative digestion in the gastrointestinal tract: diversity and evolution. Integrative and Comparative Biology 42(2): 319-326. https://doi.org/10.1093/icb/42.2.319
Matsuda, I., Murai, T., Clauss, M., Yamada, T., Tuuga, A., Bernard, H., & Higashi, S. 2011. Regurgitation and remastication in the foregut-fermenting proboscis monkey (Nasalis larvatus). Biology Letters 7(5): 786-789. https://doi.org/10.1098/rsbl.2011.0197
Perez, HG, Stevenson, CK, Lourenco, JM, & Callaway, TR 2024. Understanding rumen microbiology: an overview. Encyclopedia 4(1): 148-157. https://doi.org/10.3390/encyclopedia4010013
Pérez, W., Duro, S., & Gündemir, O. 2023. Anatomical Differences in the Omasum of Weaning Calves Fed with Different Diets. Anatomia 2(2): 176-188. https://doi.org/10.3390/anatomia2020016
Pokhrel, B., & Jiang, H. 2024. Postnatal growth and development of the rumen: integrating physiological and molecular insights. Biology 13(4): 269. https://doi.org/10.3390/biology13040269
Song, Y., Lan,
Tanabe, Shintaro. 2016. On Beef. Poplar Publishing, Tokyo. 169pp. ISBN: 9784591152461
Vendl, C., Munn, A., Leggett, K., & Clauss, M. 2017. Merycism in western gray (Macropus fuliginosus) and red kangaroos (Macropus rufus). Mammalian Biology 86: 21-26. https://doi.org/10.2307/4088763
Wiktionary contributors. 2026, January 22. Wiktionary. https://en.wiktionary.org/wiki/Wiktionary:Main_Page
Zhang, Z., Wang, Y., Si, X., Cao, Z., Li, S., & Yang, H. 2020. Rumen methanogenesis, rumen fermentation, and microbial community response to nitroethane, 2-nitroethanol, and 2-nitro-1-propanol: an in vitro study. Animals 10(3): 479. https://doi.org/10.3390/ani10030479

