Eggplant (Solanum nigrum) and nightshade (Solanum carolinense) are two species with similar names and flower shapes, so those unfamiliar with them might confuse them. However, eggplant and nightshade are completely different species. Eggplant is a cultivated plant that does not grow wild and has no thorns, while nightshade grows wild and has thorns. However, although eggplant is now thornless, its ancestral species, distributed in South Asia and Southeast Asia, did have thorns. This indicates that the thorns were removed through artificial selection. Not only that, but it is known that its morphology has also been significantly altered by humans. This can be seen as evolution to be convenient for humans, or it can be seen as eggplant "labor parasitism," relying on humans for protection, illustrating the two sides of evolution. The eggplant, thus created, was initially used as medicine mainly in Asia and the Middle East, but after selective breeding reduced its toxicity, its unripe fruit began to be widely used in cooking, including in Europe, Africa, and America. It seems to have been introduced to Japan between the Asuka and Nara periods. On the other hand, the horsetail (Solanum carolinense), native to North America, grows spontaneously, is highly prolific, and is disliked due to its numerous thorns and poison. The flowers of the eggplant and the horsetail are quite similar, and it has been found that the anthers of their stamens are only released when vibrated by a specific type of bee. This is a mechanism to increase the certainty of pollination. The fruit of the horsetail remains poisonous even when ripe, which is a seemingly mysterious phenomenon in nature. There are various theories, but research suggests that it is related to the low nutritional value of the fruit. This article will explain the classification, history, evolution, reproductive ecology, pollination ecology, and seed dispersal of the eggplant and the horsetail.
- Two species with names that include "eggplant" and similar flower shapes.
- What is the difference between eggplant and nightshade?
- Are there any other similar species?
- How did eggplants evolve? Did eggplants originally have thorns?!
- What is the history of eggplants? They weren't originally suitable for eating!?
- What are the health benefits of eggplant?
- Why is the horse nightshade disliked?
- What is the structure of the flowers of eggplant and nightshade?
- The distinctive anthers of the Solanum genus depend on the vibrations of bees!?
- Why are there both long and short pistils within the same individual?
- What is the structure of the fruit of eggplant and nightshade?
- How does the nightshade plant disperse its seeds? Why do some animals eat its poisonous fruit?
- References
- Source
Two species with names that include "eggplant" and similar flower shapes.
Eggplant (Solanum melongena), also known as eggplant, is a perennial plant believed to originate from eastern India, although there are various theories. In Japan, it is cultivated as an annual. It has been cultivated in Southeast Asia since ancient times (Kodansha, 2013). It gained popularity in the Near East and Asia, and in Japan, it was introduced from China during the Nara period (Aoba, 2013). The unripe purple fruit is mainly used for food (Kodansha, 2013), and the fruit, stem, calyx, root, and flowers were also sometimes used for medicinal purposes (Tanaka, 1995).
Solanum carolinense, also known as the "evil eggplant" or "wild eggplant," is a naturalized plant native to North America that has spread throughout the world. This perennial herb grows in cultivated fields, urban areas, wastelands, and farmlands. Known as a highly invasive weed, it was first recorded in Japan in 1906 (Meiji era) and spread from the early Showa period. Its Japanese name comes from its tendency to proliferate and be difficult to eradicate, as well as the toxicity of its fruit.
Both belong to the Solanum genus of the nightshade family, and their names and flower shapes are similar, so people who haven't seen them before might confuse the two species.
What is the difference between eggplant and nightshade?
However, eggplant and nightshade are completely different species.
Eggplants are cultivated plants and do not grow wild, but nightshade (Solanum carolinense) does grow wild.
In terms of morphology, eggplants have no thorns or stellate hairs on their stems and leaves, whereas horsetails usually have thorns and stellate hairs on their stems and leaves.
Regarding the flowers, eggplants have purple corollas, while horse nightshade (Solanum carolinense) has white to pale purple corollas.
Regarding the fruit, eggplants ripen from purple to yellowish-brown, become elongated and enlarged, and become edible when ripe due to a decrease in solanine content. In contrast, nightshade rhynchophylla ripens from green to orange-yellow, is small and spherical, and contains a large amount of solanine even when ripe, making it inedible.
While eggplant originates from India, nightshade is native to North America. Aside from belonging to the same genus, they are so different that they have almost nothing in common.








Are there any other similar species?
The genus Solanum includes many other species, but only a limited number of them are herbaceous and upright.
For those interested in learning about climbing and woody varieties of the Solanum genus, please see our other article.



How did eggplants evolve? Did eggplants originally have thorns?!
How did eggplants evolve?
While various theories have been explored over the years, a scientific method called "molecular phylogenetic analysis," which uses genes to study the evolutionary process of organisms, has now been developed, allowing us to measure the closeness of different species more accurately than before.
Similar research using the same methodology has revealed that the species Solanum insanum is the closest relative to the eggplant (Ranil et al., 2017; Aubriot et al., 2018). Therefore, examining this species may provide insights into how the eggplant evolved.
Since Solanum insanum is a recently separated species, its distribution is not yet fully understood, but it has so far been found in South Asia (Pakistan, India, Sri Lanka), Southeast Asia (Thailand, Laos, Cambodia, Myanmar, Indonesia, Philippines), China, Taiwan, and Madagascar.
From this, it is certain that South Asia and Southeast Asia are the origins of the eggplant. However, isolated distributions such as Madagascar are also observed. This is thought to be due to artificial introduction for cultivation.
What are the morphological differences between eggplant (Eggplant) and Solanum insanum ?
Solanum insanum has thorns all over the plant, its fruit is small and spherical, and its mesocarp is not spongy. In contrast, eggplant (Solanum nigrum), as mentioned above, has no thorns, its fruit is enlarged, and its mesocarp is spongy.
This means that eggplants were artificially selected from Solanum insanum to have no thorns, large fruits, and a spongy mesocarp.

If eggplants had thorns, people might get injured while growing them, and a more developed fruit and mesocarp would make them more satisfying to eat for humans.
Furthermore, unlike Solanum insanum, modern eggplants have the characteristic that even if multiple flowers bloom on a single pedicel (the part connecting the flower to the stem), only the one at the base will produce fruit (Tanaka, 1995). In other words, they have evolved to concentrate their limited resources into a single fruit.
Furthermore, the decrease in solanine is thought to have occurred during later periods of selective breeding.
To some people, these might seem like only good things. However, the absence of thorns and the reduction in solanine clearly make it harder for the plant to protect itself from predators. In fact, anyone who has grown eggplants will likely agree that they are easily eaten by pests and birds.
In other words, "eggplants evolved to specialize in fruit production, abandoning defenses, based on the premise that they would grow while being protected from predators by humans." This type of evolution is widely known in cultivated plants.
While it's true that modern eggplants lack thorns and are easier to grow, they still require constant attention. This can be seen as a testament to the efforts of ancient people, or perhaps a form of "labor parasitism" on humans. Remembering these things when growing eggplants might give you a sense of the wonders of evolution.
What is the history of eggplants? They weren't originally suitable for eating!?
Eggplants, with a total production of approximately 51 million tons in 2016, are the second most economically important fruit crop in the Solanaceae family after tomatoes (Aubriot et al., 2018). How did eggplants spread throughout the world?
As mentioned above, eggplant was artificially selected and domesticated from Solanum insanum, but this domestication may have occurred in multiple locations (Ranil et al., 2017; Aubriot et al., 2018). These regions are thought to be Southeast Asia and South Asia. Generally, the India-Burma region is considered to have been the center of domestication (Daunay & Janick, 2007). Interestingly, unlike the "mizunasu" that Japanese people eat today, eggplants before the progress of selective breeding had strong toxins such as solanine and were primarily used for medicinal purposes, and it seems that some kind of processing was necessary even if they were to be eaten.
In India, various words related to eggplant are recorded in Sanskrit texts dating back to around 300 BC (Mauryan dynasty), suggesting that it was widely used as food and medicine (Daunay & Janick, 2007). In Ayurveda, a traditional Indian and Sri Lankan system of medicine based on Sanskrit, the white fruit is recommended for diabetic patients, and the root for treating asthma. On the other hand, in the ancient Hindu scripture "Marukandeyā Purana" from the 4th century (Gupta dynasty), eggplant is listed among the undesirable. In modern Indian cuisine, eggplant dishes such as achar and beguni are well-known.
In China, it was used as a vegetable crop from an early period, and its existence is shown in the oldest botanical treatise , "Nanfang Caomuzhang, " written by Ji Han during the Western Jin dynasty (265-316), and in the agricultural treatise "Qi Min Yao Shu," written by Jia Sixie during the Northern Wei dynasty around the 6th century (420-581). In "Bencao Gangmu," a comprehensive work of Chinese herbal medicine completed in 1578, Li Shizhen states that medicinal preparations using the fruit, receptacle, roots, stems, and leaves can cure various diseases such as abscesses, intestinal bleeding, and toothaches. In modern Chinese cuisine, it is known to be used in dishes such as Yu Xiang Qiezi (fish-fragrant eggplant).
It is believed that eggplant was introduced to Japan around the 8th century, when the number of Japanese envoys to Tang China increased and relations with Tang China deepened. The oldest documented record is often said to be the "Shosoin Documents of Todai-ji Temple" (750 AD), but it already appears in the "Wooden Tablets of Prince Nagaya's House" (circa 710 AD) (Morishita, 1999). 710 AD was the year the capital was moved from Fujiwara-kyo to Heijo-kyo, marking the end of the Asuka period and the beginning of the Nara period. The "Wooden Tablets of Prince Nagaya's House" mention its use in pickled eggplant, and the "Engishiki," a legal code summarizing the detailed regulations of the Ritsuryo system compiled in 927 AD (mid-Heian period), indicates that it was eaten raw, pickled in soy sauce, pickled in rice bran, and dried. Initially, it was used as a gift for high-ranking individuals, but it became widely cultivated from around the Edo period and finally became a vegetable familiar to ordinary people. Currently, some well-known Japanese dishes include shigi-yaki, asazuke, mapo nasu, nasu dengaku, yakinasu, nasu miso stir-fry, and nibitashi.
Eggplants spread to Persia (present-day Iran) in the western part of the Eurasian continent very early, although the exact time is unknown. Ibn Sina, a scholar of the Samanid dynasty and considered one of the greatest intellectuals produced by the Islamic world, appears in his magnum opus "Canon of Medicine," completed in 1020, which combines ancient Arabic medicine with medical knowledge from Greece and India. Subsequently, it was mentioned by leading medical and botanical writers of medieval Persia, who stated that its toxicity could be reduced by salting and that the fruit could be used to neutralize bile and treat ear diseases.
Its arrival in Europe was late, and for a long time it was unknown in ancient Greece and Rome. However, with the arrival of Muslims in the 7th and 8th centuries, it spread along the Mediterranean coast and eventually reached Europe. However, it did not become popular until the 15th century (Santich & Bryant, 2008=2010). This may be due to its toxicity, but it also seems that one of the reasons is that it is originally a tropical plant and difficult to cultivate (Kodansha, 2013). In current Mediterranean cuisine, in addition to ratatouille, a French dish that is also well-known in Japan, there are eggplant jams (Turkey), baba ghanoush (Levant), ajapsandari (Georgia and Armenia), moussaka (Greece), caponata (Italy), tumbet (Spain), and pisto manchego (Spain).
From the 8th century onward, it was introduced to East Africa by Persian and Arab sailors. During the Age of Discovery (15th-17th centuries), it was transported from Spain to the Americas as one of the vegetables, and by the mid-17th century, it had been reported in Brazil.
What are the health benefits of eggplant?
Eggplant fruit is mostly water (over 931 TP3T), and is low in calories compared to other vegetables. This is one reason why it might not be highly rated, but on the other hand, it is also rich in dietary fiber, making it ideal for dieting.
Although it is sometimes said that it contains almost no vitamins, it does contain a balanced amount of vitamins such as vitamin B2 and vitamin C, as well as minerals such as potassium and calcium, albeit in relatively small amounts.
Studies that have comprehensively examined the health benefits of eggplant have cited the following examples (Fraikue, 2016; Gürbüz et al., 2018). While there are still many unknowns, such as how much eggplant is beneficial to consume and how it compares to other foods, avoiding excessive consumption will likely make it an important source of nutrients that helps the body.
- As an iron chelating agent, eggplant is strongly recommended as a food for pregnant women, breastfeeding mothers, and teenage girls in particular. The iron contained in eggplant can improve premenstrual syndrome, amenorrhea, and anemia in pregnant and postpartum women.
- Because dietary fiber has the property of absorbing harmful chemicals and toxins, it suppresses the proliferation and development of cancer cells.
- The component called nasunin (anthocyanin) has the effect of removing iron accumulated in the body, thus helping to normalize and control iron levels. In addition, nasunin protects humans from lipid peroxides.
- By removing harmful free radicals, relaxing blood vessels in the heart, and ultimately strengthening blood circulation, eggplant helps reduce cardiovascular diseases (stroke, cardiac arrest, and heart disease). Rich in dietary fiber, potassium, vitamin B6, and flavonoids, eggplant reduces the risk of these diseases.
- Ingesting it has the effect of lowering cholesterol. Chlorogenic acid acts as a major antioxidant, and the abundant dietary fiber promotes the absorption of blood cholesterol in order to produce bile in the liver.
- Potassium helps maintain electrolyte balance and assists in neutralizing sodium, which is necessary for blood pressure control; therefore, consuming potassium can help control blood pressure.
- Dietary fiber adds bulk to food, keeping you feeling full for a long time, so it can be used for weight loss.
- It keeps hair, nails, and skin healthy. The high levels of moisture, vitamins, minerals, and fats it contains improve the quality of hair, nails, and skin, and enhance general bodily functions, including resistance to skin cancer.
- High levels of alpha-glycosidase and angiotensin help absorb glucose in the body and, along with other nutrients, control type 2 diabetes. Additionally, phenol-rich extracts assist with the enzymes and glucose assimilation necessary for controlling diabetes. Furthermore, it is low in carbohydrates, and its dietary fiber helps control blood sugar levels.
- Consuming large quantities can prevent damage to brain cells. It also contains plant-based nutrients that enhance memory function and protect cell membranes.
- Because it contains minerals that humans need, such as magnesium, manganese, potassium, and copper, it promotes bone health.
- Dietary fiber flushes out waste products and harmful toxins, promoting healthy digestion.
- It contains antioxidants that protect brain cells, shield them from free radical damage, and help prevent brain tumors, while also maintaining mental health. The chlorogenic acid and phenolic substances it contains act as antiviral, antimutagenic, and antimicrobial agents, helping to supply gentle antibodies to control a healthy life.
- The dietary fiber it contains stimulates bowel movements, cleanses away harmful toxins, and prevents constipation.
Why is the horse nightshade disliked?
While eggplants have been so widely used by humans, the opposite is true of the nightshade (Solanum carolinense). As its name suggests, it's treated as a villain. Why is that?
Its reproductive capacity is high, it has spread to various environments around the world, it is difficult to eradicate, it has thorns, and the entire plant, including the fruit, is poisonous (Miyazaki et al., 2011).
Its native habitat is the United States and Canada, but it has spread to Haiti, Brazil, Bangladesh, Georgia, India, Nepal, South Korea, Japan, Croatia, Norway, and New Zealand.
The first recorded sighting of Solanum carolinense in Japan was in 1906 (Meiji era) at a ranch in Narita City, Chiba Prefecture, but it is believed to have been introduced mixed in with forage grass seeds imported from overseas. However, it does not appear that this population spread throughout the country.
After World War II (1945 onwards), horse nightshade (Solanum carolinense) was recorded in various locations, suggesting it may have been mixed in with military supplies transported by the occupying forces. In the 1990s, it became widely known as a highly damaging weed in fodder fields. As it is a highly damaging weed to corn, soybeans, and cotton in the United States, it may have originated from imported concentrated feed (feed containing high levels of nutrients such as protein, carbohydrates, and fats, but low in fiber). Currently, its distribution has spread from Hokkaido to Okinawa. However, it has been suggested that the records in Okinawa may be of a different species.
Its habitat is extremely broad, and it grows in areas disturbed by human activity, such as cultivated fields, grasslands, orchards, field edges, roadsides, vacant lots, forest edges, roadside plantings, and riverbanks.
Furthermore, it has a strong reproductive capacity. Solanum carolinense spreads vegetatively by sending out rhizomes that spread vertically and horizontally, producing new shoots from the root system and the underground base of the stem. Therefore, it can sprout from roots underground during tilling, or spread by root fragments being mixed into soil during construction or soil attached to plants. In fact, a study in Kyoto City confirmed clones at a distance of more than 1 km. It is possible that its nationwide distribution is mainly following this route. These characteristics make eradication difficult.
Furthermore, although it may be rare in Japan, as described later, it produces fruit and reproduces by seed dispersal. This is likely necessary to adapt to a new environment by exchanging certain genes with other plants.
Even these issues alone can have an impact on the landscape, but that's not the only problem. The entire plant has thorns, so there's a risk of injury when walking on paths or when manually pulling or cutting it to use the land. This also makes eradication difficult. Furthermore, it can get tangled in harvesting machinery and reduce the space available for livestock to use the grassland.
Another problem is its toxicity. It contains toxins such as solanine and saponin, which are common in the nightshade family. Solanidine, which is produced when solanine is hydrolyzed, is a neurotoxin that inhibits estrase, and if livestock consume it, it can cause symptoms of poisoning such as motor disorders, convulsions, coma, and liver damage including jaundice.
The fact that even within the same Solanum genus, there can be such a wide range of evaluations is a fascinating aspect of biological evolution.
What is the structure of the flowers of eggplant and nightshade?
Eggplants are very common plants, but you may not often see their flowers unless you grow them yourself. They flower from June to September, and the inflorescences are mostly solitary, rarely in the form of reduced racemes. There are hardly any peduncles. One to several flowers with purple corollas, 3 to 5 cm long, bloom downwards on pedicels 1 to 1.8 cm long (Tanaka, 1995). The calyx has star-shaped downy hairs and often has spines about 3 mm long on its outer surface.
Solanum carolinense blooms from June to October. Thick branches emerge from the middle of the stem, bearing 6 to 10 pale purple or white flowers, about 2 cm in diameter, in a cymose inflorescence at the tip. The corolla is shallowly divided into 5 lobes and opens into a saucer shape.
As mentioned above, eggplants have fewer flowers than horsetails in order to concentrate nutrients into a single fruit, but aside from this, the flowers of the Solanum genus, including these two species, share many structural similarities.

The distinctive anthers of the Solanum genus depend on the vibrations of bees!?
The stamens, in particular, exhibit a major characteristic common to the Solanum genus.
This is because the anthers, the pollen-containing part of the stamen, are large and conspicuously yellow, and these anthers taper to a point with a small hole. Such anthers are called "porous anthers" (Shimizu, 2001). What role do these special anthers play?
This is related to the fact that many species in the Solanum genus, including eggplant, perform a special type of pollination called "vibration pollination" (Raju & Rao, 2006). Some bees, such as bumblebees and carpenter bees, land on the large yellow anthers and then vibrate with the muscles in their wings, causing the pollen to fall out through the holes at the tips and be carried away. In this way, the Solanum species can prevent nectar robbing by carpenter bees and other species (the act of drilling holes in the calyx and directly taking the nectar without attaching pollen), and can provide them with only pollen for pollination (Ichikawa et al., 2011). It also helps to limit the visits of insects such as syrphidae and honeybees, which visit the flowers of various other plant species and make pollination uncertain.
The downward-facing flowers likely limit the insects that visit them to bees. Hanging onto downward-facing flowers requires strong legs and the ability to fly, and only certain types of bees possess these abilities.
Furthermore, since Solanum species do not produce nectar, it is thought that they have evolved to specialize in pollinating bees and thus deter butterflies and moths from visiting them.
In this way, the Solanum genus cleverly increased the reliability of pollination by limiting the types of insects that visit its plants.
Specifically, regarding Solanum carolinense, there is a record of the black bumblebee Bombus ignitus visiting in Japan (Ichikawa et al., 2011).
In Pennsylvania, USA, its native habitat, 138 out of 146 insects that visited the flowers were Bombus impatiens, a species of bumblebee (Kariyat et al., 2021). However, in Connecticut, several species of the genus Lassioglosum and Augochloropsis metallica of the family Halictidae are also reported to visit the flowers (Quesada-Aguilar et al., 2008).
Why are there both long and short pistils within the same individual?
It is known that eggplant flowers can have three types within the same plant: those with pistils longer than the pistil, those with pistils of the same length, and those with pistils that are shorter than the pistil (National Agriculture and Food Research Organization, 2006).
Because the length of the style within the pistil varies, flowers where the pistil is longer than the stamen are sometimes called "long-styled flowers," flowers where the pistil is the same length as the stamen are called "medium-styled flowers," and flowers where the pistil is shorter than the stamen are called "short-styled flowers."
Furthermore, it has been revealed that there are two types of flowers in Solanum carolinense: long-styled flowers and short-styled flowers (Quesada-Aguilar et al., 2008).
This characteristic of having different styles is called "heterostyly," and it can be seen in various plants (Watanabe, 2022).

However, even though eggplant and nightshade exhibit heterostyliosis, they differ from other plants in several ways. In other plants, heterostyliosis occurs "in separate individuals," so usually only one style length type is observed within a single individual. In contrast, in eggplant and nightshade, heterostyliosis occurs "within the same individual."
While this condition is sometimes referred to as andromonoecy, it's not entirely accurate.
Heterostyle, which occurs "in separate individuals," is generally thought to be a mechanism to prevent self-pollination.
So, what role does heterostylicity, which occurs "within an individual," play?
There are several hypotheses, but a study conducted in Connecticut, USA, on Solanum carolinense supports the idea that by changing the length of the pistil in this way, each flower is responsible for dividing the roles of male and female.
In other words, in long-styled flowers, the pistil is longer than the stamens, so the pistil protrudes. This means the pistil gets in the way, preventing bees from effectively pollinating by vibration and thus preventing them from collecting pollen from the stamens. However, when a bee clings to the flower, it can receive pollen from another individual. Therefore, long-styled flowers take on the role of female flowers and produce fruit.
On the other hand, in short-styled flowers, the pistil is shorter than the stamens, so the pistil is buried within the stamens and cannot be pollinated. Instead, there are no obstacles for the bee to move, so it can efficiently vibrate the flower and drop the pollen onto its body. Therefore, short-styled flowers take on the role of the male bee, and are dedicated to spreading pollen to other individuals.
Interestingly, it has been found that this effect is only exerted when a species of bumblebee , Bombus impatiens, visits.
In Connecticut, it is known that other bees of the family Halicidae also visit the flowers of Solanum carolinense. However, Bombus impatiens touches the flower more frequently and more often with shorter vibrations than other Halicidae bees, efficiently dispersing pollen. This is likely why the difference in pollen dispersal efficiency between long-styled and short-styled flowers is so clearly reflected in the numbers.
I mentioned that eggplant and nightshade flowers come in three types, but it's important to know that they don't always have all three types; some plants only have one type, and there's a lot of individual variation.
This suggests that there are situations where it is beneficial to induce heterostyly growth, and situations where it is not, depending on the type of bee that visits. It is possible that the presence of bumblebees in the environment is closely linked to the occurrence of heterostyly growth "within the individual plant."
As research into the effects of these pollinating insects on plants progresses, we will likely gain a more general understanding of why some plants have heterostyly structured stigmas while others do not.
What is the structure of the fruit of eggplant and nightshade?
The fruits of the Solanum genus are all berries.
Eggplants are classified as berries, but unlike tomatoes, which belong to the same genus, they are not sweet. They are black to purple in color, turning yellowish-brown when fully ripe. Their shape and size vary greatly depending on the variety, but most are over 6 cm in diameter. The seeds are lenticular and yellowish.
The berries of the horsetail plant are spherical, about 1.5 cm in diameter, and ripen from green to yellow.
Eggplant fruits have evolved to be large enough to be eaten, but what is their structure like?
The eggplant fruit has a nested structure consisting of the exocarp, endocarp, partition, placenta, and core (Wang et al., 2021; Nakamura, 2021).
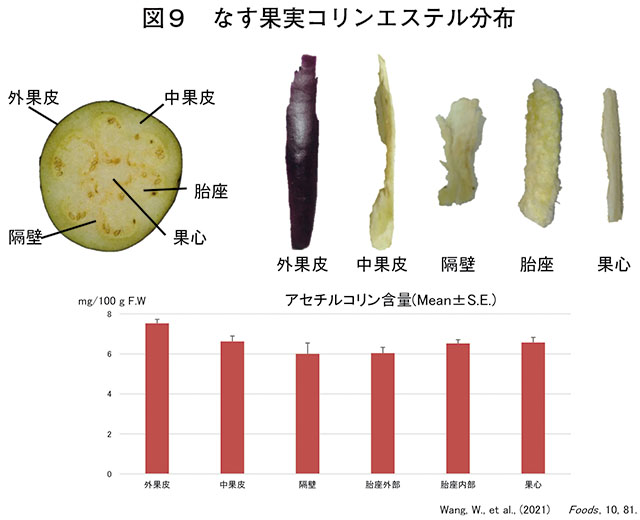
The outermost purple part of an edible, unripe fruit is called the "exocarp." The exocarp contains about four types of anthocyanins, 90% of which is nasunin, a monoacyl anthocyanin, and is the main component that creates the purple color (Ito et al., 2013).
The inside is entirely white and spongy, making it difficult to accurately identify with the naked eye.
Inside the outer pericarp is a thick, fleshy endocarp. Inside the endocarp are multiple placentas, separated by septa, and each placenta contains numerous seeds. These multiple placentas surround the core of the fruit, known as the "pith."
How does the nightshade plant disperse its seeds? Why do some animals eat its poisonous fruit?
It's safe to assume that eggplant seeds are now dispersed by human hands, as mentioned above.
How does the horse nightshade disperse its seeds?
While there are no observations that specifically record the types of animals that eat the fruit, studies in North America suggest that birds and mammals eat the fruit (Cipollini & Levey, 1997a). However, in Japan, there has been an experiment where the fruit of Solanum carolinense was placed and observed with an infrared camera, but no animals that eat it were identified (Miyazaki et al., 2011). However, I have seen Solanum carolinense growing in isolation in a planted area along a road. This could be because the roots of Solanum carolinense were mixed in with the transplanted soil, but it is also possible that the fruit is being eaten by birds.
By the way, there's a big question about the fruit of the horsetail plant. As mentioned above, the fruit of the horsetail plant contains solanine even when ripe and is poisonous. This isn't true for all plants in the Solanum genus; often, the solanine disappears when the plant ripens. So why do such unpalatable fruits exist even when ripe?
This inexplicable phenomenon has been studied in detail (Cipollini & Levey, 1997a; b).
The first possibility to consider is that the toxicity may be limiting which animals come to the fruit. This fruit is toxic to humans, and it is known that ingestion by livestock can cause motor disorders, convulsions, coma, and jaundice. On the other hand, it may be harmless to birds and some wild mammals that digest only the pulp without destroying the seeds, or because their metabolism is different. In this case, it would be possible to get only specific animals to eat the fruit and disperse the seeds. This type of hypothesis is called the "directed toxicity hypothesis."
To verify this, four species of frugivorous birds common in North America—the Japanese robin (Turdus migratorius), the bobtail quail (Colinus virginianus), the mammals Peromyscus maniculatus and Didelphis virginiana— were actually given the fruits of Solanum carolinense (a type of nightshade) along with the non-toxic fruits of Solanum americanum, and their feeding rates were compared.
As a result, all mammals and birds disliked the fruit of Solanum carolinense, which is more poisonous than Solanum nigrum, and their consumption rates were significantly lower. This suggests that the possibility of it being non-toxic to only certain animals, as suggested by the direct toxicity hypothesis, is unlikely.
If that's the case, what other reasons could there be? There are two more hypotheses that consider the relationship with the nutritional value of the fruit.
The first hypothesis is the "Removal Rate Hypothesis." If there are few birds or mammals in the habitat, or if the fruit is relatively less nutritious than other plants and therefore unpopular, the fruit will remain attached to the plant indefinitely. In that case, the fruit is more likely to be damaged by pests or fungi. If that is the case, the benefit of repelling these harmful creatures with its toxicity outweighs the disadvantage of not being eaten by animals that contribute to seed dispersal.
Conversely, if the habitat has many birds and mammals, or if the fruit is relatively more nutritious and popular than other plants, the fruit will be quickly eaten by birds and mammals, eliminating the need to protect it from pests and fungi. In this case, reducing toxicity would have the advantage of being eaten by animals that contribute to seed dispersal, outweighing the disadvantage of not being able to repel harmful pests and fungi.
Simply put, if the rate at which the fruit is eaten is low, the toxicity will be high, and if the rate at which it is eaten is high, the toxicity will be low.
The second hypothesis is the "Nutrient-Toxin Titration Hypothesis," which presents a contrasting perspective. If a plant is highly nutritious and popular with birds and mammals, it may be able to invest in increased toxicity to protect itself from pests and fungi. On the other hand, if a plant is low in nutritional value and unpopular with birds and mammals, it may not attract them at all unless its toxicity is also low.
In short, this hypothesis states that if the rate at which the fruit is eaten is high, the toxicity will be high, and if the rate at which it is eaten is low, the toxicity will be low.
Both arguments seem convincing, but this can only be confirmed by actually conducting an experiment.
This was also tested in the same study that investigated the direct toxicity hypothesis. The results showed that the fruit of the highly toxic Solanum carolinense was not particularly nutritious. This supports the "removal rate hypothesis."
In summary, due to factors such as the scarcity of birds and mammals in its habitat, and the relatively low nutritional value of its fruit compared to other plants making it unpopular, the carnation fruit needed to be protected from pests and fungi for extended periods, and even if it was disliked by birds and mammals, it increased its toxicity.
As mentioned above, the horsetail plant is specialized to grow in barren land, so it may have been difficult for it to sufficiently increase the nutritional value of its fruit. Therefore, as a last resort, it likely protects its fruit with glycoalkaloids such as solanine. Although it is sometimes treated as a villain because it is poisonous, knowing these circumstances on the part of the horsetail plant gives it a different perspective.
References
Aubriot, X., Knapp, S., Syfert, MM, Poczai, P., & Buerki, S. 2018. Shedding new light on the origin and spread of the brinjal eggplant (Solanum melongena L.) and its wild relatives. American Journal of Botany 105(7): 1175-1187. https://doi.org/10.1002/ajb2.1133
Aoba, Takashi. 2013. Encyclopedia of Japanese Vegetable Culture History. Yasaka Shobo, Tokyo. 486pp. ISBN: 9784896941609
Cipollini, M. L., & Levey, D. J. 1997a. Why are some fruits toxic? Glycoalkaloids in solanumand fruit choice by vertebrates. Ecology 78(3): 782-798. https://doi.org/10.1890/0012-9658(1997)078[0782:WASFTG]2.0.CO;2
Cipollini, M. L., & Levey, D. J. 1997b. Antifungal activity of Solanum fruit glycoalkaloids: implications for frugivory and seed dispersal. Ecology 78(3): 799-809. https://doi.org/10.1890/0012-9658(1997)078[0799:AAOSFG]2.0.CO;2
Daunay, MC, & Janick, J. 2007. History and iconography of eggplant. Chronica Horticulturae 47(3): 16-22. ISSN: 0578-039X, https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=79218125888314fab4f5275db48d5625b124c915
Fraikue, FB 2016. Unveiling the potential utility of eggplant: A review. Conference Proceedings of INCEDI 1: 883-895. https://www.researchgate.net/publication/310846026
Gürbüz, N., Uluişik, S., Frary, A., Frary, A., & Doğanlar, S. 2018. Health benefits and bioactive compounds of eggplant. Food Chemistry 268: 602-610. https://doi.org/10.1016/j.foodchem.2018.06.093
Ichikawa, S., Kurahashi, T., & Ikuru, S. 2011. Possibility of a novel pollination mode by flower-visiting bees and yellow-breasted bumblebees collected in Kagawa Prefecture. Bulletin of the Faculty of Agriculture, Kagawa University 63(116): 43-59. ISSN: 0368-5128, http://id.nii.ac.jp/1731/00003553
Ito, K., Akutsu, T., Oyama, T., Watanabe, T., Yamazaki, K., Kakuzaki, F., & Uda, Y. 2013. Preparation of antioxidant powder containing nasunin and chlorogenic acid from eggplant pre-pickling liquid. Journal of the Japanese Society for Food Science and Technology 60(1): 30-37. https://doi.org/10.3136/nskkk.60.30
Kariyat, RR, Bentley, TG, Nihranz, CT, Stephenson, AG, De Moraes, CM, & Mescher, MC 2021. Inbreeding in Solanum carolinense alters floral attractants and rewards and adversely affects pollinator visitation. American Journal of Botany 108(1): 74-82. https://doi.org/10.1002/ajb2.1594
Kodansha. 2013. The Book of Vegetables: Seasonal Ingredients Gentle on the Body. Kodansha, Tokyo. 255pp. ISBN: 9784062183420
Miyazaki, Kei; Nishida, Tomoko; and Urakawa, Shuji. 2011. Weed Monograph 6. Solanum carolinense L.. Weed Research 56(3): 154-165. https://doi.org/10.3719/weed.56.154
Morishita, Masahiro. 1999. Fruit characteristics and history of 'Mizunasu' (water eggplant). Research Report of Osaka Prefectural Agricultural and Forestry Technology Center 35: 34-39. https://agriknowledge.affrc.go.jp/RN/2010691355
Nakamura, K. 2021. Food functions of eggplant and functional foods. Vegetable Information 211: 40-48. ISSN: 2436-9586, https://vegetable.alic.go.jp/yasaijoho/senmon/2110_chosa1.html
National Agriculture and Food Research Organization. 2006. Encyclopedia of the Latest Agricultural Technology. Rural Culture Association, Tokyo. v, 2003pp. ISBN: 9784540051630, https://lib.ruralnet.or.jp/nrpd/#koumoku=13429
Quesada-Aguilar, A., Kalisz, S., & Ashman, TL 2008. Flower morphology and pollinator dynamics in Solanum carolinense (Solanaceae): implications for the evolution of andromonoecy. American Journal of Botany 95(8): 974-984. https://doi.org/10.3732/ajb.0800106
Raju, AS, & Rao, SP 2006. Nesting habits, floral resources and foraging ecology of large carpenter bees (
Ranil, RH, Prohens, J., Aubriot, X., Niran, HML, Plazas, M., Fonseka, RM, … & Knapp, S. 2017. Solanum insanum L.(subgenus Leptostemonum Bitter, Solanaceae), the neglected wild progenitor of eggplant (S. melongena L.): a review of taxonomy, characteristics and uses aimed at its enhancement for improved eggplant breeding. Genetic Resources and Crop Evolution 64: 1707-1722. https://doi.org/10.1007/s10722-016-0467-z
Santich, B. & Bryant, G. 2008. Edible: An illustrated guide to the world's food plants. National Geographic, Washington, DC. 360pp. ISBN: 9781426203725 [= 2010. Illustrated Guide to the Culture of Edible Plants of the World: Origin, History, Distribution, Cultivation, and Cuisine. Shufusha, Tokyo. 360pp. ISBN: 9784903530352]
Shimizu, Takemi. 2001. Illustrated Dictionary of Botanical Terms. Yasaka Shobo, Tokyo. xii, 323pp. ISBN: 9784896944792
Tanaka, Koji. 1995. Herbal Health Methods: Effectiveness and Usage at a Glance. Kodansha, Tokyo. 123pp. ISBN: 9784061953727
Wang, W., Yamaguchi, S., Suzuki, A., Wagu, N., Koyama, M., Takahashi, A., … & Nakamura, K. 2021. Investigation of the distribution and content of acetylcholine, a novel functional compound in eggplant. Foods 10(1): 81. https://doi.org/10.3390/foods10010081
Watanabe, Kenta. 2022. Current Ecological and Evolutionary Biological Perspectives on "Heterostyle." Okinawa National College of Technology Bulletin 16: 31-45. https://doi.org/10.51104/nitokinawacollege.16.0_31
Source
This article is a significantly expanded version of one included in the following book.

