
Arisaema ringens, Arisaema serratum, and Arisaema serratum are representative species of the Arisaema genus. The Arisaema genus is extremely similar in shape, making it a difficult group to distinguish. To put it bluntly, distinguishing between species in the Arisaema genus is extremely difficult. There is considerable variation in classification between regional populations, and the classification changes frequently depending on what is considered a species, making it difficult for an amateur to judge. However, in the case of Arisaema ringens, Arisaema serratum, and Arisaema serratum, it is possible to distinguish them by observing the spathe. The spathe is a highly specialized morphology unique to the Araceae family, but recent research has surprisingly revealed that pollination occurs by trapping fungus gnats inside until they die. Furthermore, active research in Japan has shown that the types of fungus gnats differ for each Arisaema species, preventing competition and hybridization. The bright red, ripe fruits of the Arisaema genus are remarkably similar and common to all species, and seed dispersal by birds remains the same. However, some species have a summer fruiting season, suggesting the possibility of seed dispersal specifically tailored to bulbuls. This article will explain the classification, pollination ecology, and seed dispersal of the Arisaema genus.
- Extremely similar Arisaema species found locally in Japan
- What are the differences between Arisaema ringens, Arisaema serratum, and Arisaema serratum?
- What is the structure of a flower in the Arisaema genus?
- Fungus gnats are trapped inside Arisaema flowers until they die!?
- Different species of Arisaema plants attract different types of fungus gnats!?
- The fruit is a berry, and the seeds are probably dispersed only by birds?
- It seems that there are two groups of Arisaema species whose fruits ripen in summer and others whose fruits ripen in autumn or winter!?
- References
- Source
Extremely similar Arisaema species found locally in Japan
Arisaema limbatum, also known as ear-shaped ahniant
Arisaema aequinoctiale, also known as the red bellflower, is a perennial herb distributed only in Honshu (Kanto, Chubu, Hiroshima, and Yamaguchi prefectures) and Shikoku in Japan, growing in evergreen broad-leaved forests near the coast.
Arisaema japonicum, also known as jack-in-the-pulpit, is a perennial herb distributed in Shikoku and Kyushu in Japan (up to the Tokara Islands) and in Korea (Ulleungdo), growing in forest undergrowth and forest edges from lowlands to mountainous areas.
All of these plants belong to the genus Arisaema in the Araceae family and are remarkably similar in shape. A distinctive feature that distinguishes them from other plants in this genus is that their stems are underground rhizomes, and they have a flattened, spherical corm. A "pseudostem" grows on top of this, and "sheath-like leaves" are attached to wrap around the pseudostem. Photosynthesis is carried out by the "ordinary leaves" that grow at the end of the pseudostem, which are palmately compound, whornately compound, or rarely simple. In the case of compound leaves, the separated parts are called "leaflets." Each leaf has one axillary bud.

The Arisaema genus is known for containing calcium oxalate crystals in its plants, making it toxic to mammals, and for its ability to change sex (Kono, 1988).
What are the differences between Arisaema ringens, Arisaema serratum, and Arisaema serratum?
To be frank, distinguishing between species of the Arisaema genus is extremely difficult. There is considerable variation in classification among regional populations, and the classification frequently changes depending on what is considered a species, requiring the publication of new books each time (Murata, 2015; 2018). In the latest literature, the Arisaema genus in Japan is divided into 53 species (Murata, 2018).
Therefore, it would be difficult for an amateur to easily distinguish between them.
Here, we will consider the differences between relatively well-known species such as Arisaema ringens, Arisaema serratum, and Arisaema serratum. For more detailed information, please refer to books on the subject.
These three species differ in that, firstly, in Arisaema ringens and Arisaema serratum, the inflorescence stalk is longer than the pseudostem (at least in females), there are 5 to 7 or more leaflets, and the average number of ovules per ovary is 10 or more, whereas in Arisaema serratum, the inflorescence stalk is shorter than the pseudostem, there are 7 or more leaflets, and the average number of ovules per ovary is 8 or less. Also, the opening of the spathe in Arisaema serratum does not spread out in an ear-like shape.
The difference between Arisaema ringens and Arisaema serratum is that in Arisaema ringens, the opening of the spathe is broad and ear-shaped, with a spreading width of 8 mm or more, while in Arisaema serratum, the opening of the spathe is only slightly ear-shaped, with a spreading width of less than 8 mm.
Also, although it's not mentioned in the identification key in the field guide, in Arisaema ringens, the inflorescence appendage is rod-shaped to somewhat club-shaped with a swollen, rounded tip that extends completely outside the spathe opening, while in Arisaema serratum, the inflorescence appendage is rod-shaped and only slightly extends from the spathe opening. I think this point is relatively easy to understand.
However, Arisaema ringens and Arisaema serratum share many similarities within the Arisaema genus, such as having a chromosome number of 2n=26 and having 9 to 17 leaflets in females, making identification between the two species difficult.
In any case, it's difficult to identify it accurately without the flowers in bloom.

















What is the structure of a flower in the Arisaema genus?
The Arisaema genus has flowers that are significantly different from other plants, and it has many specialized terms.
In most varieties, the flowers bloom from spring to early summer.
First, the flowers of the Arisaema genus are enclosed in a structure called a "spathe." This is a common structure in the Araceae family, where the bracts are large, developed leaves, and the inflorescence can be seen inside.
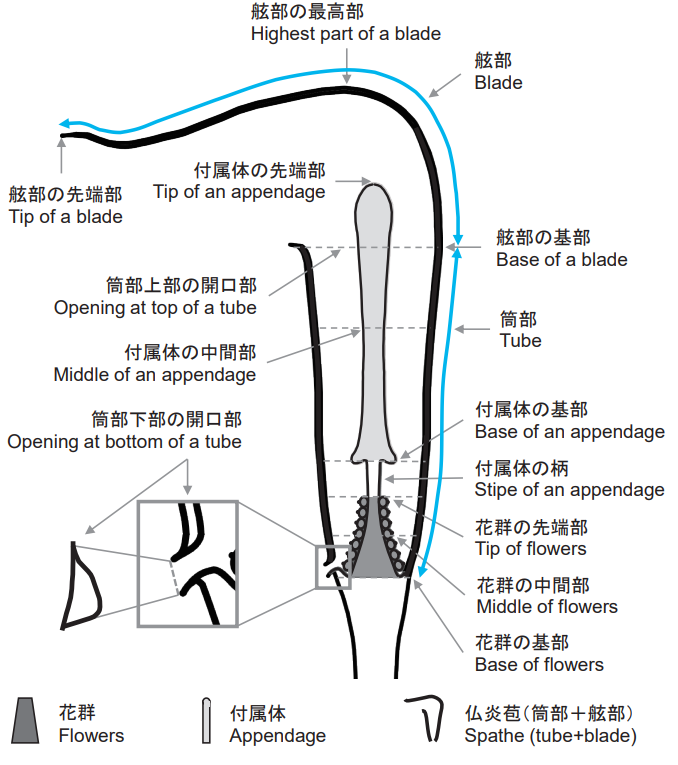
The spathe is connected to the pseudostem by a "peduncles," and structurally it can be divided into a "tube" that directly encloses the inflorescence, a "mouth" that serves as the entrance to the inflorescence, and a "limb" that loosely closes the mouth in a roof-like shape.
The inflorescences of Araceae plants are also specialized and are called "spadixes." In a spadix, the flower axis is thickened and fleshy, and numerous sessile flowers are densely packed around it. Spadixes have an "appendage" attached to them, which grows upward and can sometimes be seen from the tube of the spathe.
The Arisaema genus is dioecious, meaning it has separate male and female plants. Male plants have male flowers on their spadixes, while female plants have female flowers on their spadixes.
Interestingly, the sex of Arisaema species is determined by their nutritional status. Smaller plants become male, capable of producing gametes even with relatively few resources, while larger plants that receive sufficient nutrients and grow into larger specimens change sex to become female, producing female or hermaphroditic flowers. Such unique dioecious plants are sometimes called "pseudodioecious."
Arisaema ringens flowers from March in warmer regions and around May in the Tohoku region. The opening of the spathe is broad and ear-shaped, with a width of 8 mm or more at the opening.
Arisaema ringens flowers from March to April, and the opening of its spathe is slightly ear-shaped, with a width of less than 8 mm.
The Arisaema serratum blooms from March to April, and the opening of its spathe is only slightly open.

Fungus gnats are trapped inside Arisaema flowers until they die!?
The flowers of the Arisaema genus are extremely specialized, so I'm very curious about what kind of insects visit and pollinate them.
This phenomenon has been studied by various researchers. As a result, it has been discovered that very small flies, such as those belonging to the families Mycetophilidae and Sciaridae, are visiting the area (Tanaka & Hirano, 2000; Vogel & Martens, 2000; Suetsugu, 2022).
These flies have a unique life cycle, feeding on mold and fungal mycelium during their larval stage, and are well known to be abundant in forests. Considering that Arisaema species inhabit forests, they can be considered excellent pollinators.
Specifically, how do fungus gnats get to flowers and pollinate Arisaema flowers?
First, fungus gnats are attracted to the spathe using the inflorescence appendage as a guide.
Unfortunately, it is still not well understood how fungus gnats are attracted to spathes. In Arisaema sikokianum, it is thought that fungus gnats are attracted to the flower by the scent that mimics a mushroom and emits a mushroom-like odor. However, no such scent components have been identified in other Arisaema species. Nevertheless, it has been suggested that fungus gnats may emit sex pheromones that are undetectable to the human nose.
Next, fungus gnats attracted to the spathe penetrate the interior and reach the spadix.
However, the inside of this spathe is smooth and also has a rat-proof barrier, so fungus gnats attracted by the scent fall in and cannot get out from the top. As a result, the fungus gnats are left to roam freely inside the spathe.
If the fallen flower is from a male plant, there is an exit at the bottom of the spathe. When it emerges, it will be covered in pollen from the male flower, but this does not pose a threat to the fungus gnats' lives.
The problem arises with female plants. Unlike male plants, female plants do not have an exit at the bottom! While pollination can occur on the Arisaema side by fungus gnats thrashing around inside the spathe, just like with male plants, the fungus gnats simply die.
In other words, the fungus gnats are trapped twice, unable to escape, and then ultimately killed. This could be considered a very cruel method of pollination.
While the exact reasons for killing the fungus gnats are not yet fully understood, one theory suggests that the fungus gnats' continued thrashing until death contributes to efficient pollination.
Different species of Arisaema plants attract different types of fungus gnats!?
While the pollination method of the Arisaema genus is certainly unique, there are some puzzling aspects. The Arisaema genus is diverse, and different species can sometimes be found in the same location. In such cases, since they utilize the same fungus gnats as pollinators, do problems like competition or hybridization not occur?
In relation to this point, research has been conducted in Japan (Matsumoto et al., 2021). This study investigated the specific types of fungus gnats that visit five species of plants distributed at the Okayama Ski Resort: Arisaema maekawae, Arisaema angustatum, Arisaema peninsulae, Arisaema pseudoangustatum var. pseudoangustatum, and Arisaema ovale.
As a result, it was discovered that the types of fungus gnats attracted differ depending on the type of Arisaema plant.
Specifically, the most frequently observed species were Megophthalmidia sp. in Arisaema umbellata, Cordyla sp. in Arisaema cordata, Mycetophila sp. (a species of fungus gnat) in Arisaema serratum, Trichosia sp. in Arisaema japonica, and Anatella sp. in Arisaema serratum.
This fact suggests that Arisaema species prevent competition and hybridization by attracting other types of fungus gnats to their flowers.
Other studies have shown that the types of fungus gnats that visit different species differ between Arisaema sikokianum and Arisaema tosaense in Otaki-Okawa Prefectural Natural Park, Kagawa Prefecture (Matsumoto et al., 2019), and in Tenkawa Village, Nara Prefecture, the types of fungus gnats that visit different species differ between Arisaema cucullatum , Arisaema kishidae, and Arisaema yamatense ( Kakishima et al., 2020). There are studies that have shown that the types of fungus gnats that visit different species of nikoense differ (Maeda & Takahashi, 2021), and studies that have shown that the types of fungus gnats that visit different species of Arisaema nagiense and Arisaema ovale also differ (Matsumoto et al., 2023).
The specific mechanism by which different fungus gnat species are attracted is not yet understood. However, as mentioned above, if the fungus gnats emit sex pheromones that are undetectable to the human nose, then differences in these pheromones may be the cause of these differences. In any case, this is undoubtedly an interesting result that illustrates the relationship between plant speciation and insects.
The fruit is a berry, and the seeds are probably dispersed only by birds?
The fruit, like all Arisaema species, is a berry, ovoid in shape. Since it lacks a pedicel, it grows densely, resembling an aggregate fruit. When ripe, it changes from green to vermilion and contains a small number of seeds. The seeds are nearly spherical, milky white, and occasionally spotted.
As evidenced by the fact that the fruit turns red when ripe, it is clear that the fruit is designed to attract animals, suggesting that seed dispersal relies on animals.
However, not all animals eat the fruit; it is primarily birds that eat the fruit and disperse the seeds. In Japanese research, mammals have rarely been observed eating this fruit, and the only animal that does eat the fruit, the field mouse, also eats the seeds, so it is thought not to contribute to seed dispersal (Oishi et al., 2020).
The reason mammals do not eat the fruits of the Arisaema genus is thought to be because the fruits contain calcium oxalate crystals, which are toxic to mammals. However, there is evidence that Himalayan black bears and American black bears have eaten them overseas, so some believe that Japanese black bears may also eat them (Maeda and Takahashi, 2021).
It seems that there are two groups of Arisaema species whose fruits ripen in summer and others whose fruits ripen in autumn or winter!?
Although there are differences in the number of fruits that grow on an inflorescence, the fruits of the Arisaema genus are basically very similar among species, to the point that they are indistinguishable when viewed individually. However, it has been found that there are interestingly two types of fruiting periods in the Arisaema genus (Kobayashi, 2017; Kobayashi et al., 2017).
For most of the 39 groups in the genus Arisaema, the fruiting period is from mid-September to January, during autumn and winter. However, the fruiting period for the five groups of Arisaema undulatifolium (excluding Arisaema undulatifolium subsp. undulatifolium, including Arisaema nambae, Arisaema minus, Arisaema undulatifolium ssp. uwajimense), and Arisaema seppikoense is from late June to August, during summer.
Do these ecological differences also lead to differences in the types of birds that are responsible for seed dispersal?
To investigate this matter, surveys using automated cameras are being conducted in western Japan (Hyogo, Okayama, and Hiroshima prefectures).
As a result, in this study, among the seven groups of plants whose fruiting season is from autumn to winter (Arisaema serratum, Arisaema serratum, Arisaema serratum, Arisaema urashima, Arisaema urashima, and Arisaema ringens), the pale thrush, copper pheasant, and brown-eared bulbul were found to be feeding on the fruit. In another study, conducted in southern Kanto (Hadano, Kanagawa Prefecture), research on Arisaema serratum confirmed that the brown-eared bulbul, red-flanked blue robin, pale thrush, copper pheasant, and red-billed leiothrix were feeding on the fruit. Similar results were shown in a study in Ishikawa Prefecture (Oishi et al., 2020). Of these, the copper pheasant is considered a seed-eater that destroys seeds inside its body, so it is highly likely that it does not contribute to seed dispersal. The fruit disappears slowly.
On the other hand, in the four summer groups whose fruiting period is from late June to August (Arisaema takahashiense, Arisaema harimanum, Arisaema ringens, and Arisaema seppiforme), a different trend was observed where bulbuls and pheasants consumed the fruit. In another study in southern Kanto, only bulbuls consumed the fruit of Arisaema ringens. As mentioned above, it is highly likely that pheasants do not contribute to seed dispersal, so in effect, bulbuls are the only ones responsible for seed dispersal. The fruit disappears rapidly.
In other words, while a variety of birds eat the fruit of Arisaema species that bear fruit in autumn and winter, only a very limited number of birds, such as the brown-eared bulbul, eat the fruit of Arisaema species that bear fruit in summer.

So why were these differences observed?
Since most species in the Arisaema genus have a fruiting period from autumn to winter, it is likely that the fruiting period for the Arisaema genus was originally from autumn to winter.
This is the time of year when various migratory birds arrive in the Japanese archipelago as winter visitors, seeking fruit to feed on. Some birds also migrate south within Japan, like vagrant birds. Other plants, in sync with this migration, begin to bear fruit.
Therefore, it is thought that the Arisaema genus efficiently disperses its seeds not only through resident birds but also through these wintering and migratory birds.
However, some species of Arisaema serratum and Arisaema tetrandra have evolved to produce fruit in the summer. The reason for this is not yet fully understood, but strong interaction with bulbuls may have spurred this evolution.
The brown-eared bulbul is a resident bird and has an extremely large population in Japan compared to other bird species. Therefore, in environments where brown-eared bulbuls are numerous, fruit is likely to be scarce during the summer (July-August) breeding season when there are fewer fruit-bearing plant species.
During such times, the fruits of the Arisaema genus, which bear conspicuous aggregate fruits on the forest floor visible from a distance, likely become an ideal food source for brown-eared bulbuls.
As a result, a group emerged that specialized for bulbuls and bore fruit in the summer. Bulbuls consumed most of the fruit, and a higher percentage of the fruit was carried away compared to the group that bore fruit in autumn and winter, which may have allowed the seeds to be efficiently dispersed into diverse environments. This was the case with some of the Arisaema serratum and Arisaema tetrandra groups.
Furthermore, female plants in the summer fruiting group tend to have longer peduncles than petioles. This may also be an evolutionary adaptation to elongate the peduncle and attract bulbuls to eat the fruit.
The fact that plants change in close relation to Japanese animals, not only in terms of pollinating insects but also in terms of fruiting period, teaches us that we should not consider these living things in isolation.
References
Kakishima, S., Sueyoshi, M., & Okuyama, Y. 2020. Floral visitors of critically endangered Arisaema cucullatum (Araceae) endemic to Kinki region of Japan. Bulletin of the National Museum of Nature and Science. Series B, Botany 46(1): 47-53. https://www.kahaku.go.jp/albums/abm.php?d=4697&f=abm00001696.pdf&n=L_BNMNS_B46-1_47.pdf
Kobayashi, Yoshiki. 2017. Thirty Years with Arisaema: What We Have Learned from Surveys and Research on the Arisaema Group. Classification 17(2): 113-127. https://doi.org/10.18942/bunrui.01702-02
Kobayashi, Yoshiki; Kitamura, Shunpei; and Murata, J. 2017. Differentiation of fruit maturation and seed dispersal by birds in Japanese Arisaema species (Araceae). Journal of Japanese Plant Research 92(4): 199-213. https://doi.org/10.51033/jjapbot.92_4_10799
Kono, Shoichi. 1988. The World of Plants: An Invitation to Natural History, No. 2. Kyoikusha, Tokyo. 143pp. ISBN: 9784315506631
Natsuki Maeda and Kazuaki Takahashi. 2021. Flowering, fruiting, pollination, and seed dispersal of Arisaema species: A case study from Mt. Asama. Nagano University Bulletin 42(3): 23-57. http://id.nii.ac.jp/1025/00001331/
Matsumoto, TK, Miyazaki, Y., Sueyoshi, M., Senda, Y., Yamada, K., & Hirobe, M. 2019. Pre-pollination barriers between two sympatric Arisaema species in northern Shikoku Island, Japan. American Journal of Botany 106(12): 1612-1621. https://doi.org/10.1002/ajb2.1389
Matsumoto, TK, Hirobe, M., Sueyoshi, M., & Miyazaki, Y. 2021. Selective pollination by fungus gnats potentially functions as an alternative reproductive isolation among five Arisaema species. Annals of Botany 127(5): 633-644. ISSN: 0305-7364, https://doi.org/10.1093/aob/mcaa204
Matsumoto, TK, Sueyoshi, M., Sakata, S., Miyazaki, Y., & Hirobe, M. 2023. Two closely related species of the Arisaema ovale group (Araceae) selectively attract male fungus gnats of different Anatella species (Diptera: Mycetophilidae). Plant Systematics and Evolution 309(1): 4. https://doi.org/10.1007/s00606-022-01839-7
Murata, J. 2015. Araceae. In: Ohashi, H., Kadota, Y., & Kihara, H. (Eds.), Wild Plants of Japan (Revised New Edition), Vol. 1, pp. 93-100. Heibonsha. ISBN: 9784582535310
Murata, J., Ohno, J., Kobayashi, Y., and Toma, T. 2018. Illustrated Guide to Arisaema species found in Japan. Hokuryukan, Tokyo. 360pp. ISBN: 9784832610057
Oishi, Rihoko; Maeda, Taisei; and Kitamura, Shunpei. 2020. The effectiveness of birds as seed dispersers of Arisaema serratum (Araceae) in temperate forests of Japan: Effects on fruit removal and germination. Bird Research 16: A1-A14. https://doi.org/10.11211/birdresearch.16.A1
Suetsugu, K. 2022. Arisaema: Pollination by lethal attraction. Plants, People, Planet 4(3): 196-200. https://doi.org/10.1002/ppp3.10261
Tanaka, Hajime & Hirano, Takahisa. 2000. The Face of Flowers: Wisdom for Bearing Fruit. Yama-kei Publishers, Tokyo. 191pp. ISBN: 9784635063043
Vogel, S., & Martens, J. 2000. A survey of the function of the lethal kettle traps of Arisaema (Araceae), with records of pollinating fungus gnats from Nepal. Botanical Journal of the Linnean Society 133(1): 61-100. ISSN: 0024-4074, https://doi.org/10.1006/bojl.1999.0317
Source
This article is a significantly expanded version of a piece originally published in the following book.

